

Unusual accelerated rate of deletions and insertions in toxin genes in the venom glands of the pygmy copperhead (Austrelaps labialis) from Kangaroo island
- PMID: 18307759
- PMCID: PMC2287176
- DOI: 10.1186/1471-2148-8-70
Unusual accelerated rate of deletions and insertions in toxin genes in the venom glands of the pygmy copperhead (Austrelaps labialis) from Kangaroo island
Abstract
Background: Toxin profiling helps in cataloguing the toxin present in the venom as well as in searching for novel toxins. The former helps in understanding potential pharmacological profile of the venom and evolution of toxins, while the latter contributes to understanding of novel mechanisms of toxicity and provide new research tools or prototypes of therapeutic agents.
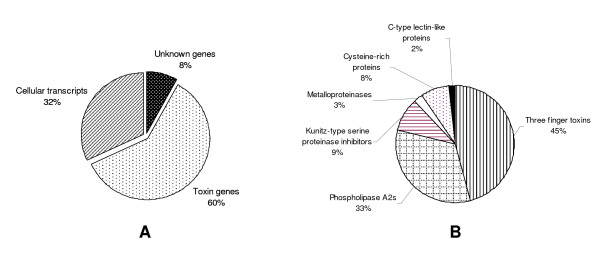
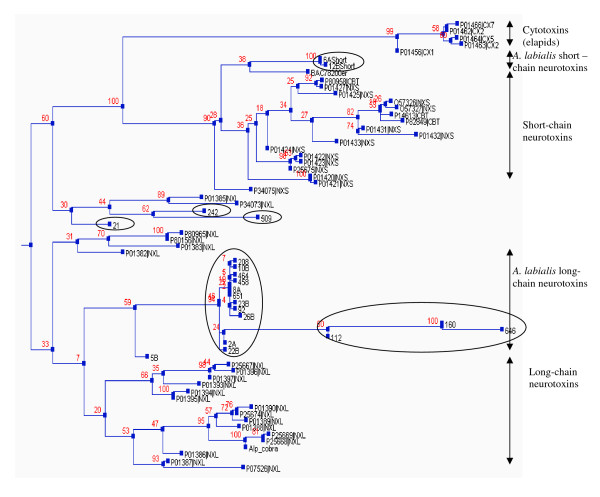
Results: The pygmy copperhead (Austrelaps labialis) is one of the less studied species. In this present study, an attempt has been made to describe the toxin profile of A. labialis from Kangaroo Island using the cDNA library of its venom glands. We sequenced 658 clones which represent the common families of toxin genes present in snake venom. They include (a) putative long-chain and short-chain neurotoxins, (b) phospholipase A2, (c) Kunitz-type protease inhibitor, (d) CRISPs, (e) C-type lectins and (f) Metalloproteases. In addition, we have also identified a novel protein with two Kunitz-type domains in tandem similar to bikunin.
Conclusion: Interestingly, the cDNA library reveals that most of the toxin families (17 out of 43 toxin genes; approximately 40%) have truncated transcripts due to insertion or deletion of nucleotides. These truncated products might not be functionally active proteins. However, cellular transcripts from the same venom glands are not affected. This unusual higher rate of deletion and insertion of nucleotide in toxin genes may be responsible for the lower toxicity of A. labialis venom of Kangroo Island and have significant effect on evolution of toxin genes.
Figures


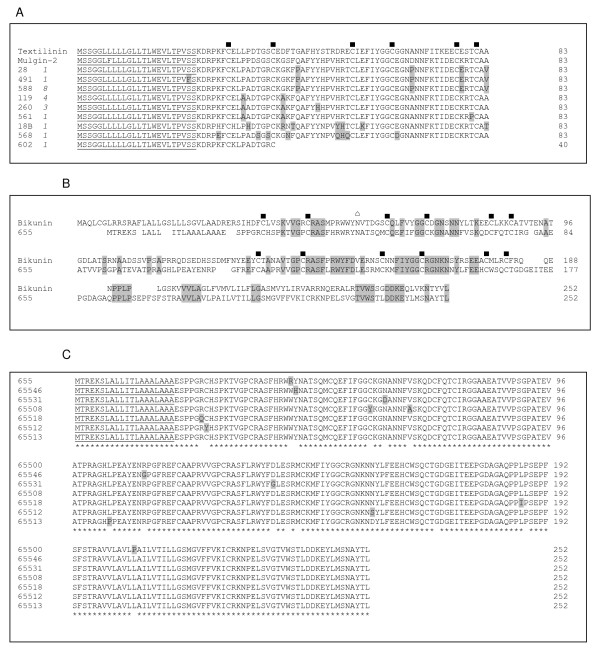
 , N-glycosylation site. Conserved residues are highlighted. C) Alignment of isoforms of two Kunitz-type domains. The presence of two Kunitz-type domains was further confirmed by PCR using gene specific primer and reverse primer from cDNA construction kit (for details, see Materials and methods). *, consensus residues and residues that are different from the consensus sequence are highlighted. The number of clones is shown in italics and the predicted signal peptide using SignalP 3.0 is underlined.
, N-glycosylation site. Conserved residues are highlighted. C) Alignment of isoforms of two Kunitz-type domains. The presence of two Kunitz-type domains was further confirmed by PCR using gene specific primer and reverse primer from cDNA construction kit (for details, see Materials and methods). *, consensus residues and residues that are different from the consensus sequence are highlighted. The number of clones is shown in italics and the predicted signal peptide using SignalP 3.0 is underlined.

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
