

Regulation of the epithelial Na+ channel by the protein kinase CK2
- PMID: 18308722
- PMCID: PMC3259572
- DOI: 10.1074/jbc.M704532200
Regulation of the epithelial Na+ channel by the protein kinase CK2
Abstract
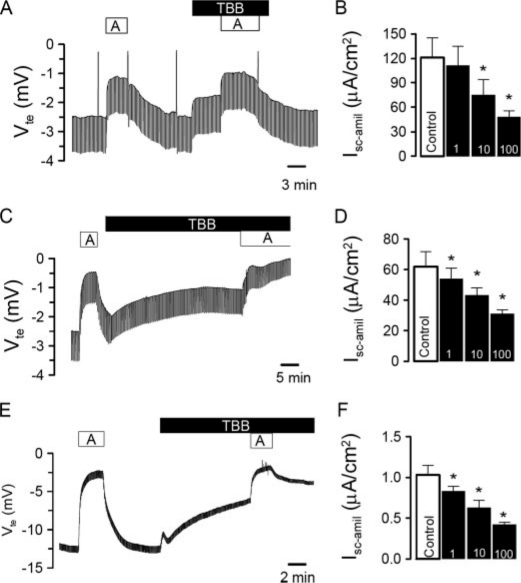
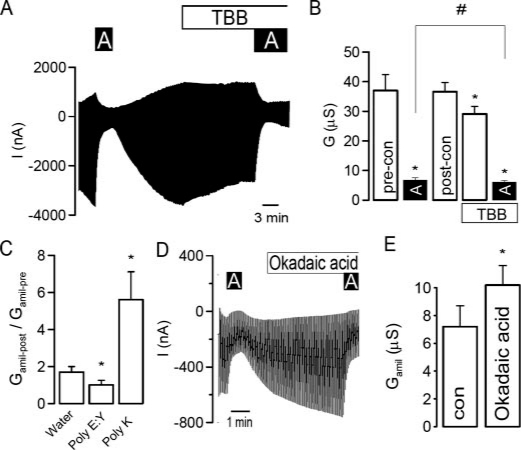
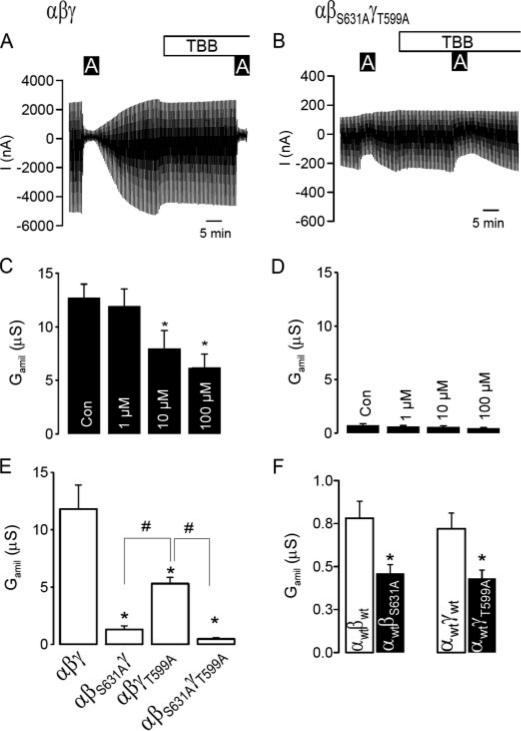
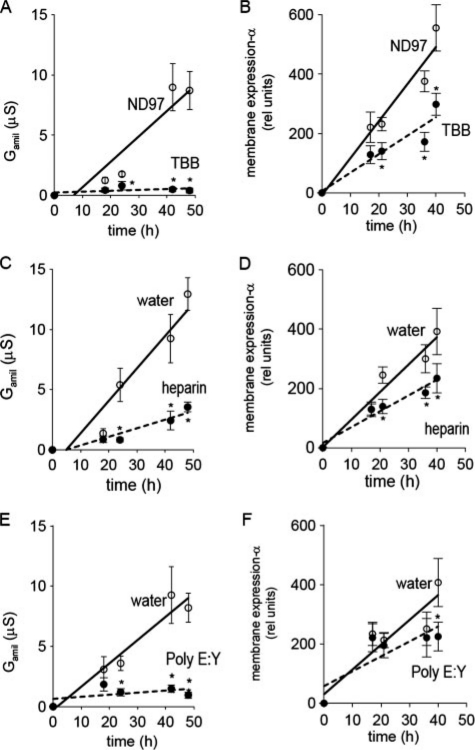
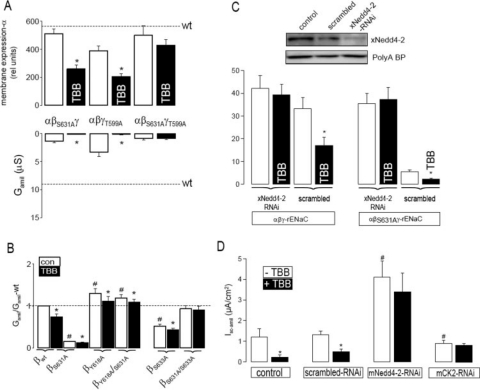
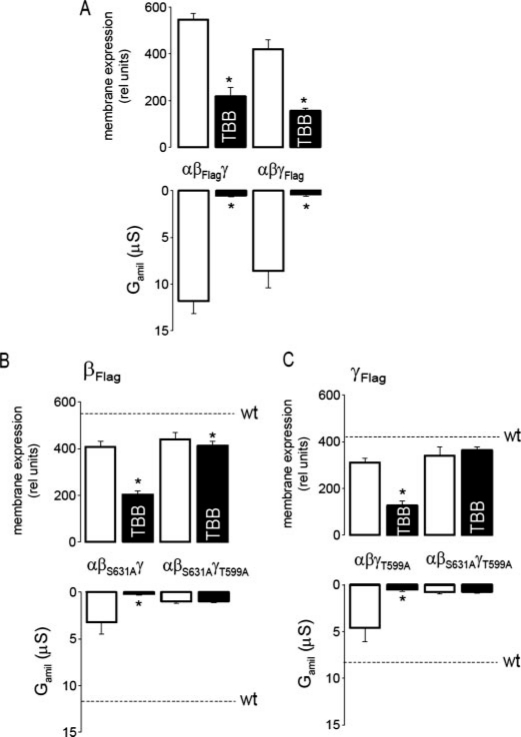
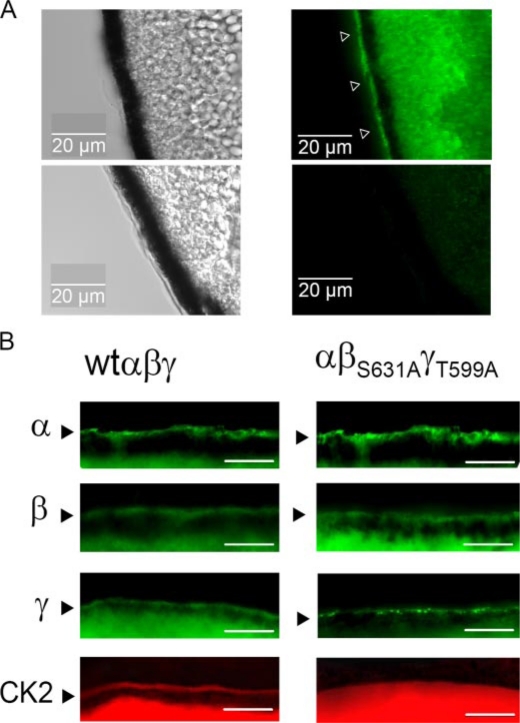
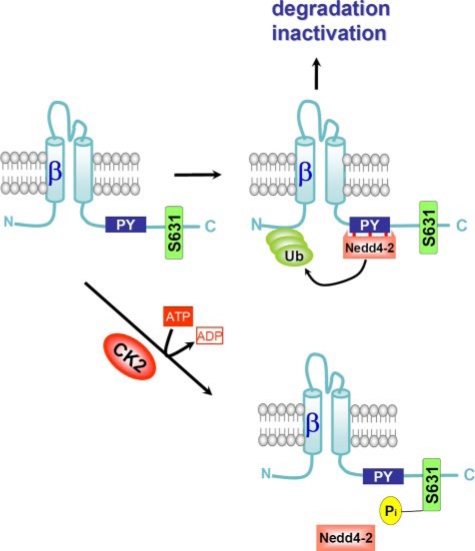
CK2 is a ubiquitous, pleiotropic, and constitutively active Ser/Thr protein kinase that controls protein expression, cell signaling, and ion channel activity. Phosphorylation sites for CK2 are located in the C terminus of both beta- and gamma-subunits of the epithelial Na(+) channel (ENaC). We examined the role of CK2 on the regulation of both endogenous ENaC in native murine epithelia and in Xenopus oocytes expressing rENaC. In Ussing chamber experiments with mouse airways, colon, and cultured M1-collecting duct cells, amiloride-sensitive Na(+) transport was inhibited dose-dependently by the selective CK2 inhibitor 4,5,6,7-tetrabromobenzotriazole (TBB). In oocytes, ENaC currents were also inhibited by TBB and by the structurally unrelated inhibitors heparin and poly(E:Y). Expression of a trimeric channel lacking both CK2 sites (alphabeta(S631A)gamma(T599A)) produced a largely attenuated amiloride-sensitive whole cell conductance and rendered the mutant channel insensitive to CK2. In Xenopus oocytes, CK2 was translocated to the cell membrane upon expression of wt-ENaC but not of alphabeta(S631A)gamma(T599A)-ENaC. Phosphorylation by CK2 is essential for ENaC activation, and to a lesser degree, it also controls membrane expression of alphabetagamma-ENaC. Channels lacking the Nedd4-2 binding motif in beta-ENaC (R561X, Y618A) no longer required the CK2 site for channel activity and siRNA-knockdown of Nedd4-2 eliminated the effects of TBB. This implies a role for CK2 in inhibiting the Nedd4-2 pathway. We propose that the C terminus of beta-ENaC is targeted by this essential, conserved pleiotropic kinase that directs its constitutive activity toward many cellular protein complexes.
Figures
References
-
- Bachhuber, T., König, J., Voelcker, T., Mürle, B., Schreiber, R., and Kunzelmann, K. (2005) J. Biol. Chem. 280 31587-31594 - PubMed
-
- Becchetti, A., Malik, B., Yue, G., Duchatelle, P., Al Khalili, O., Kleyman, T. R., and Eaton, D. C. (2002) Am. J. Physiol. Renal Physiol. 283 F1030-F1045 - PubMed
-
- Bildl, W., Strassmaier, T., Thurm, H., Andersen, J., Eble, S., Oliver, D., Knipper, M., Mann, M., Schulte, U., Adelman, J. P., and Fakler, B. (2004) Neuron 43 847-858 - PubMed
-
- Cai, Y., Anyatonwu, G., Okuhara, D., Lee, K. B., Yu, Z., Onoe, T., Mei, C. L., Qian, Q., Geng, L., Wiztgall, R., Ehrlich, B. E., and Somlo, S. (2004) J. Biol. Chem. 279 19987-19995 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
