

Metabotropic glutamate receptors activate dendritic calcium waves and TRPM channels which drive rhythmic respiratory patterns in mice
- PMID: 18308826
- PMCID: PMC2479557
- DOI: 10.1113/jphysiol.2007.149021
Metabotropic glutamate receptors activate dendritic calcium waves and TRPM channels which drive rhythmic respiratory patterns in mice
Abstract
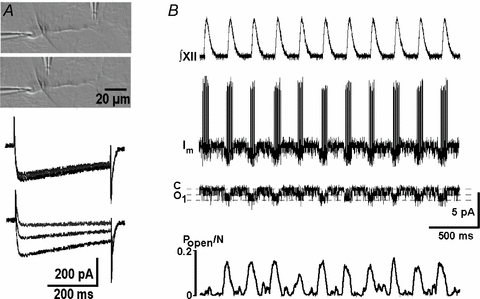
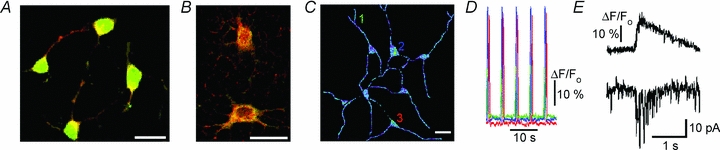
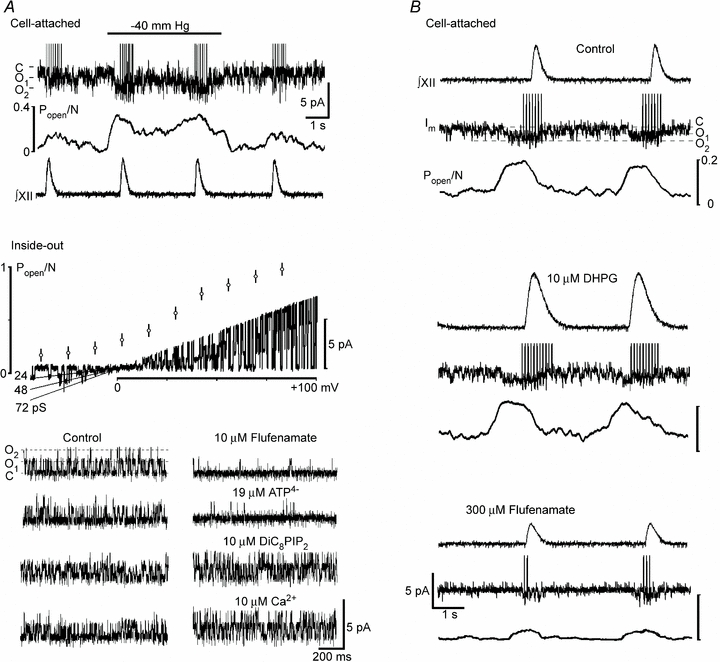
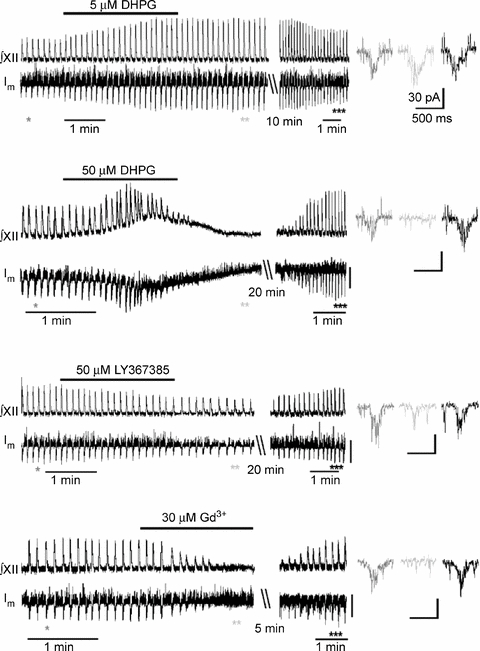
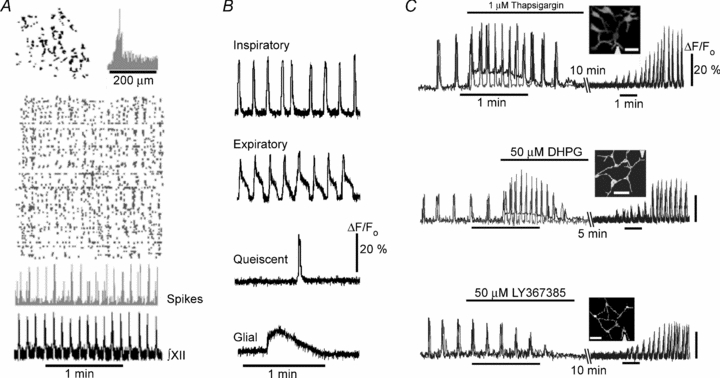
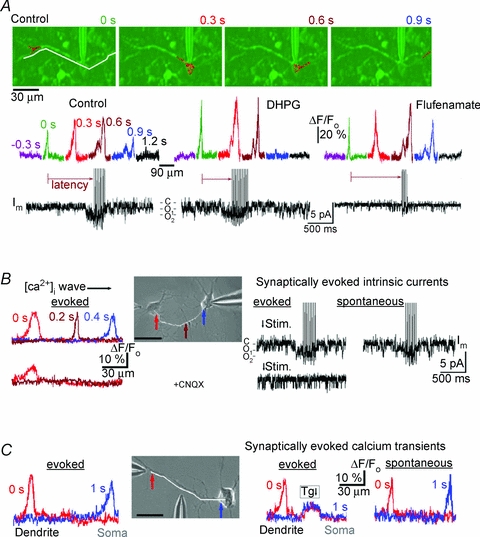
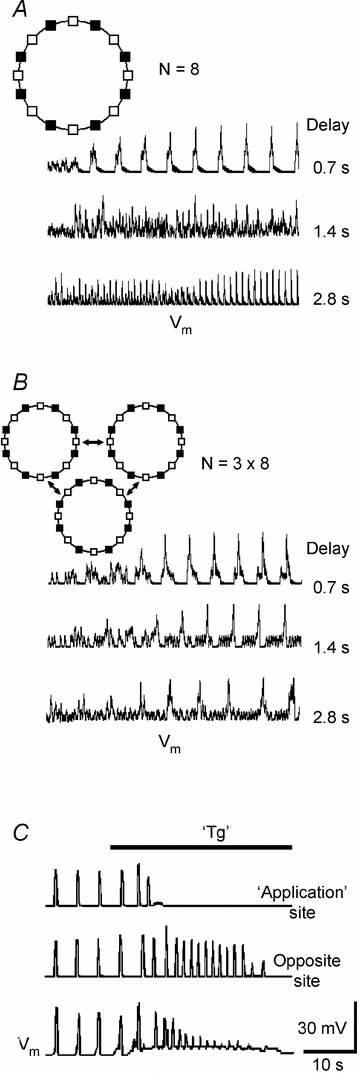
Respiration in vertebrates is generated by a compact network which is located in the lower brainstem but cellular mechanisms which underlie persistent oscillatory activity of the respiratory network are yet unknown. Using two-photon imaging and patch-clamp recordings in functional brainstem preparations of mice containing pre-Bötzinger complex (preBötC), we examined the actions of metabotropic glutamate receptors (mGluR1/5) on the respiratory patterns. The agonist DHPG potentiated and antagonist LY367385 depressed respiration-related activities. In the inspiratory neurons, we observed rhythmic activation of non-selective channels which had a conductance of 24 pS. Their activity was enhanced with membrane depolarization and after elevation of calcium from the cytoplasmic side of the membrane. They were activated by a non-hydrolysable PIP(2) analogue and blocked by flufenamate, ATP4- and Gd3+. All these properties correspond well to those of TRPM4 channels. Calcium imaging of functional slices revealed rhythmic transients in small clusters of neurons present in a network. Calcium transients in the soma were preceded by the waves in dendrites which were dependent on mGluR activation. Initiation and propagation of waves required calcium influx and calcium release from internal stores. Calcium waves activated TPRM4-like channels in the soma and promoted generation of inspiratory bursts. Simulations of activity of neurons communicated via dendritic calcium waves showed emerging activity within neuronal clusters and its synchronization between the clusters. The experimental and theoretical data provide a subcellular basis for a recently proposed group-pacemaker hypothesis and describe a novel mechanism of rhythm generation in neuronal networks.
Figures
Comment in
-
A 'group pacemaker' mechanism for respiratory rhythm generation.J Physiol. 2008 May 1;586(9):2245-6. doi: 10.1113/jphysiol.2008.153627. J Physiol. 2008. PMID: 18450782 Free PMC article. No abstract available.
Similar articles
-
A 'group pacemaker' mechanism for respiratory rhythm generation.J Physiol. 2008 May 1;586(9):2245-6. doi: 10.1113/jphysiol.2008.153627. J Physiol. 2008. PMID: 18450782 Free PMC article. No abstract available.
-
Metabotropic glutamate receptor activity induces a novel oscillatory pattern in neonatal rat hypoglossal motoneurones.J Physiol. 2005 Feb 15;563(Pt 1):139-59. doi: 10.1113/jphysiol.2004.079509. Epub 2004 Dec 20. J Physiol. 2005. PMID: 15611018 Free PMC article.
-
Metabotropic glutamate receptors (mGluR5) activate transient receptor potential canonical channels to improve the regularity of the respiratory rhythm generated by the pre-Bötzinger complex in mice.Eur J Neurosci. 2012 Jun;35(11):1725-37. doi: 10.1111/j.1460-9568.2012.08091.x. Epub 2012 May 22. Eur J Neurosci. 2012. PMID: 22612431 Free PMC article.
-
Tuning and playing a motor rhythm: how metabotropic glutamate receptors orchestrate generation of motor patterns in the mammalian central nervous system.J Physiol. 2006 Apr 15;572(Pt 2):323-34. doi: 10.1113/jphysiol.2005.100610. Epub 2006 Feb 9. J Physiol. 2006. PMID: 16469790 Free PMC article. Review.
-
Neuronal mechanisms of respiratory rhythm generation: an approach using in vitro preparation.Jpn J Physiol. 1997 Oct;47(5):385-403. doi: 10.2170/jjphysiol.47.385. Jpn J Physiol. 1997. PMID: 9504127 Review.
Cited by
-
Calcium-activated nonspecific cation current and synaptic depression promote network-dependent burst oscillations.Proc Natl Acad Sci U S A. 2009 Feb 24;106(8):2939-44. doi: 10.1073/pnas.0808776106. Epub 2009 Feb 5. Proc Natl Acad Sci U S A. 2009. PMID: 19196976 Free PMC article.
-
Dendritic A-Current in Rhythmically Active PreBötzinger Complex Neurons in Organotypic Cultures from Newborn Mice.J Neurosci. 2018 Mar 21;38(12):3039-3049. doi: 10.1523/JNEUROSCI.3342-17.2018. Epub 2018 Feb 19. J Neurosci. 2018. PMID: 29459371 Free PMC article.
-
Breathing matters.Nat Rev Neurosci. 2018 Jun;19(6):351-367. doi: 10.1038/s41583-018-0003-6. Nat Rev Neurosci. 2018. PMID: 29740175 Free PMC article. Review.
-
Effects of eugenol on respiratory burst generation in newborn rat brainstem-spinal cord preparations.Pflugers Arch. 2018 Feb;470(2):385-394. doi: 10.1007/s00424-017-2074-z. Epub 2017 Sep 29. Pflugers Arch. 2018. PMID: 28963585
-
From circuits to behaviour: motor networks in vertebrates.Curr Opin Neurobiol. 2010 Feb;20(1):116-25. doi: 10.1016/j.conb.2010.01.002. Epub 2010 Feb 6. Curr Opin Neurobiol. 2010. PMID: 20138753 Free PMC article. Review.
References
-
- Augustine GJ, Santamaria F, Tanaka K. Local calcium signaling in neurons. Neuron. 2003;40:331–346. - PubMed
-
- Baker RE, Ballantyne D, Bingmann D, Jones D, Widman G. Rhythm generation in medullary cultures of newborn rats. Int J Dev Neurosci. 1995;13:799–809. - PubMed
-
- Busselberg D, Bischoff AM, Richter DW. A combined blockade of glycine and Ca2+-dependent K+ channels abolishes respiratory rhythm. Neuroscience. 2003;122:831–841. - PubMed
-
- Butera RJ, Rinzel J, Smith JC. Models of respiratory rhythm generation in the pre-Botzinger complex. I. Bursting pacemaker neurons. J Neurophysiol. 1999;82:382–397. - PubMed
-
- Campbell CG, Spray DC, Wolkoff AW. Extracellular ATP4− modulates organic anion transport by rat hepatocytes. J Biol Chem. 1993;268:15399–15404. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources