

Wall teichoic acids are dispensable for anchoring the PNAG exopolysaccharide to the Staphylococcus aureus cell surface
- PMID: 18310032
- PMCID: PMC2292800
- DOI: 10.1099/mic.0.2007/013292-0
Wall teichoic acids are dispensable for anchoring the PNAG exopolysaccharide to the Staphylococcus aureus cell surface
Abstract
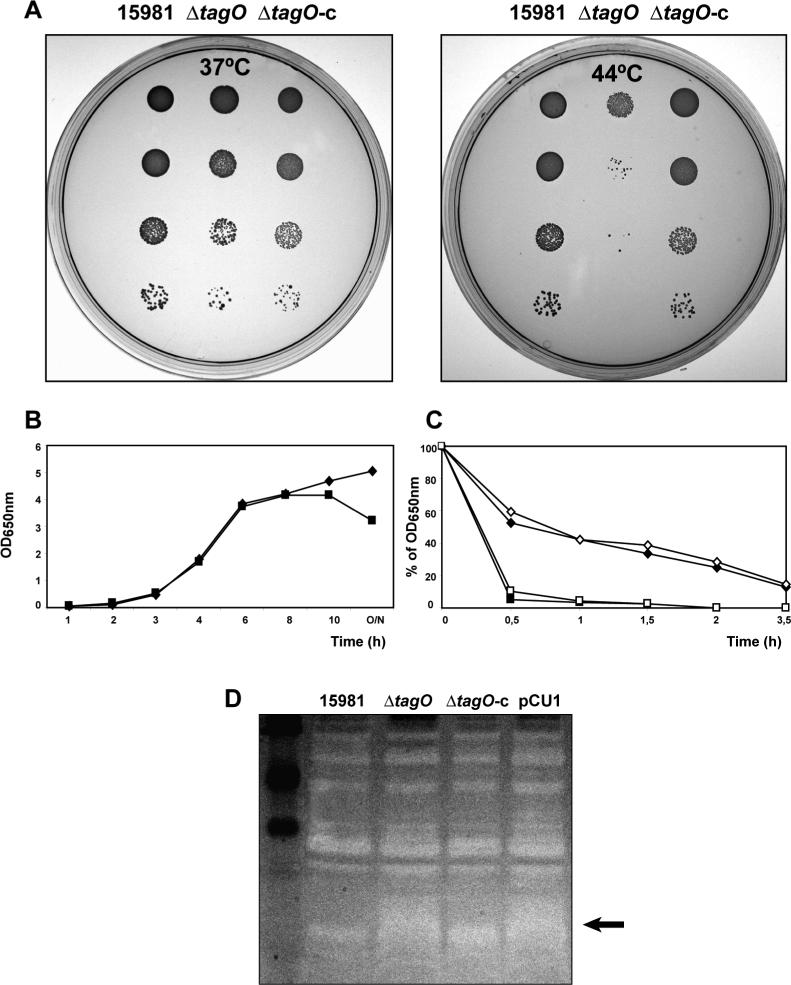
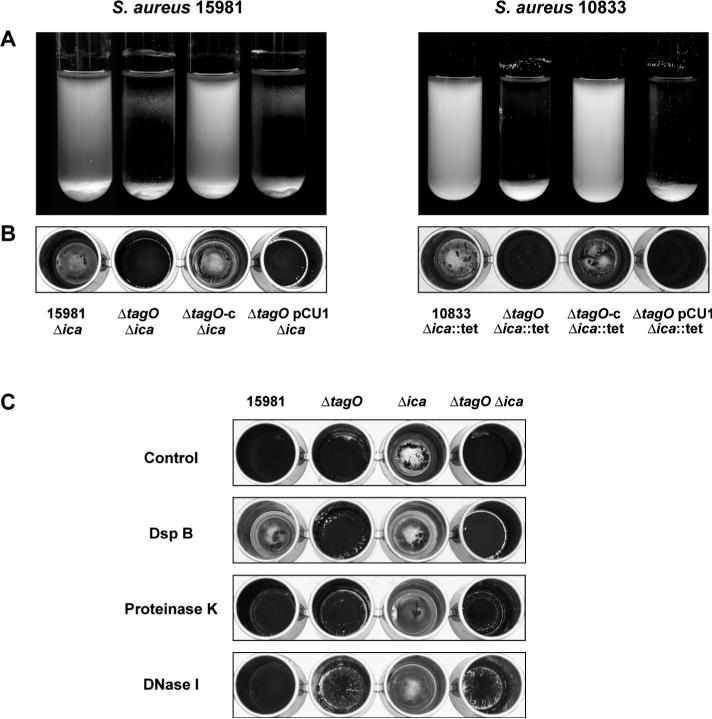
Biofilm formation in Staphylococcus aureus is usually associated with the production of the poly-N-acetylglucosamine (PNAG) exopolysaccharide, synthesized by proteins encoded by the icaADBC operon. PNAG is a linear beta-(1-6)-linked N-acetylglucosaminoglycan that has to be partially deacetylated and consequently positively charged in order to be associated with bacterial cell surfaces. Here, we investigated whether attachment of PNAG to bacterial surfaces is mediated by ionic interactions with the negative charge of wall teichoic acids (WTAs), which represent the most abundant polyanions of the Gram-positive bacterial envelope. We generated WTA-deficient mutants by in-frame deletion of the tagO gene in two genetically unrelated S. aureus strains. The DeltatagO mutants were more sensitive to high temperatures, showed a higher degree of cell aggregation, had reduced initial adherence to abiotic surfaces and had a reduced capacity to form biofilms under both steady-state and flow conditions. However, the levels as well as the strength of the PNAG interaction with the bacterial cell surface were similar between DeltatagO mutants and their corresponding wild-type strains. Furthermore, double DeltatagO DeltaicaADBC mutants displayed a similar aggregative phenotype to that of single DeltatagO mutants, indicating that PNAG is not responsible for the aggregative behaviour observed in DeltatagO mutants. Overall, the absence of WTAs in S. aureus had little effect on PNAG production or anchoring to the cell surface, but did affect the biofilm-forming capacity, cell aggregative behaviour and the temperature sensitivity/stability of S. aureus.
Figures





References
-
- Aly R, Shinefield HR, Litz C, Maibach HI. Role of teichoic acid in the binding of Staphylococcus aureus to nasal epithelial cells. J Infect Dis. 1980;141:463–465. - PubMed
-
- Augustin J, Rosenstein R, Wieland B, Schneider U, Schnell N, Engelke G, Entian KD, Gotz F. Genetic analysis of epidermin biosynthetic genes and epidermin-negative mutants of Staphylococcus epidermidis. Eur J Biochem. 1992;204:1149–1154. - PubMed
-
- Baddiley J, Buchanan JG, Hardy FE, Martin RO, Rajbhandary UL, Sanderson AR. The structure of the ribitol teichoic acid of Staphylococcus aureus H. Biochimica et biophysica acta. 1961;52:406–407. - PubMed
-
- Baddiley J. Teichoic acids in bacterial coaggregation. Microbiology. 2000;146(Pt 6):1257–1258. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources