

A newly identified extrinsic input triggers a distinct gastric mill rhythm via activation of modulatory projection neurons
- PMID: 18310125
- PMCID: PMC2430018
- DOI: 10.1242/jeb.015222
A newly identified extrinsic input triggers a distinct gastric mill rhythm via activation of modulatory projection neurons
Abstract
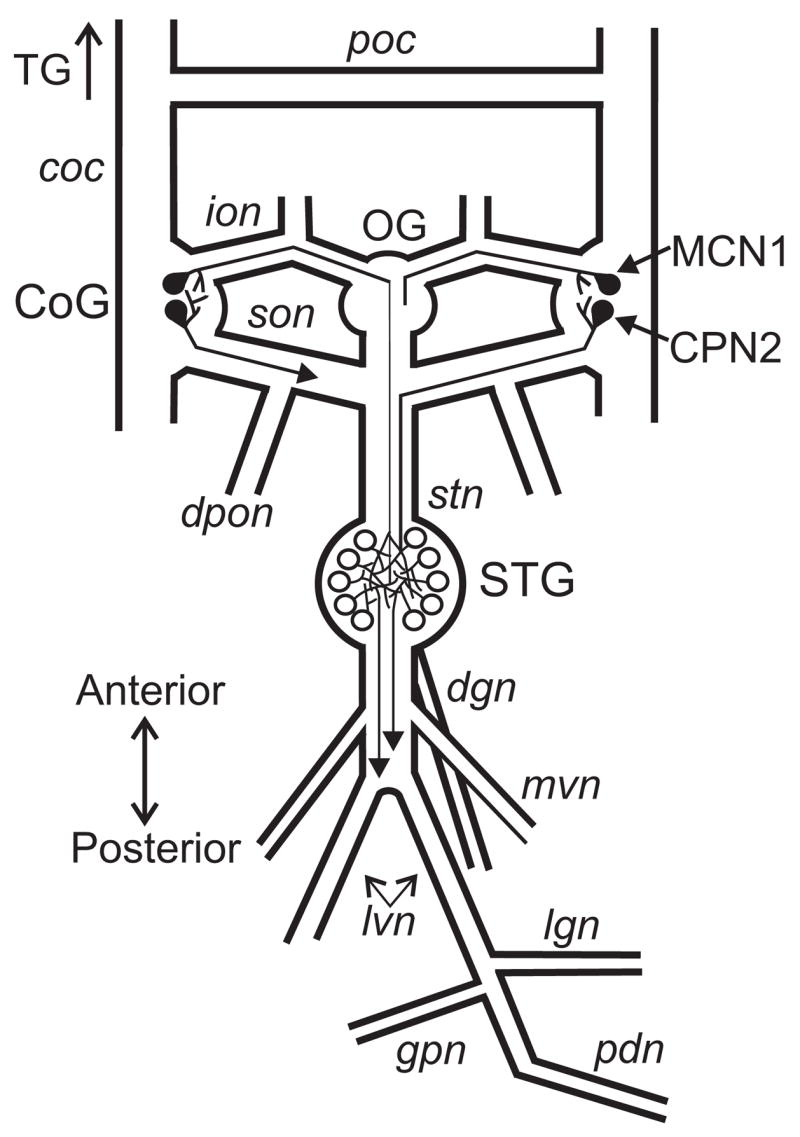
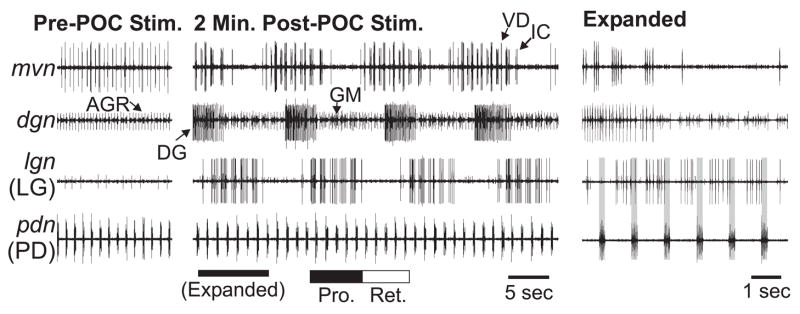
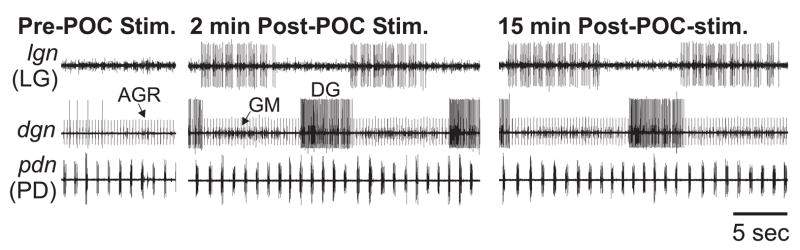
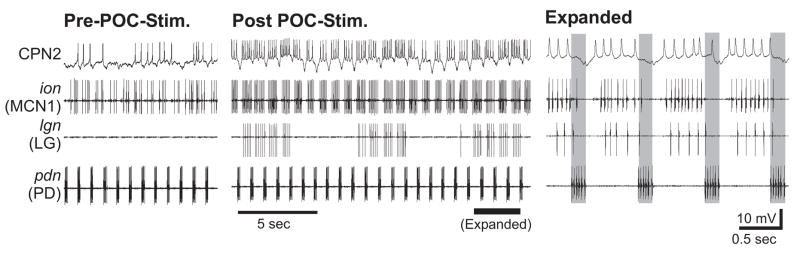
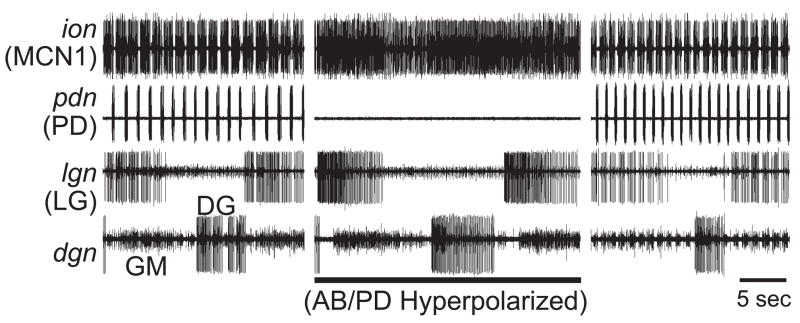
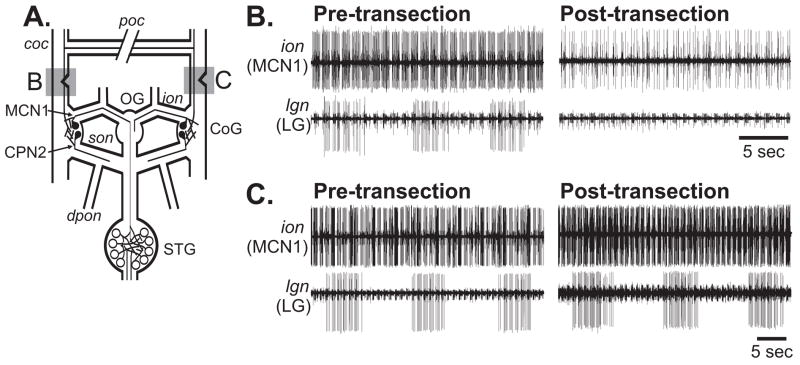
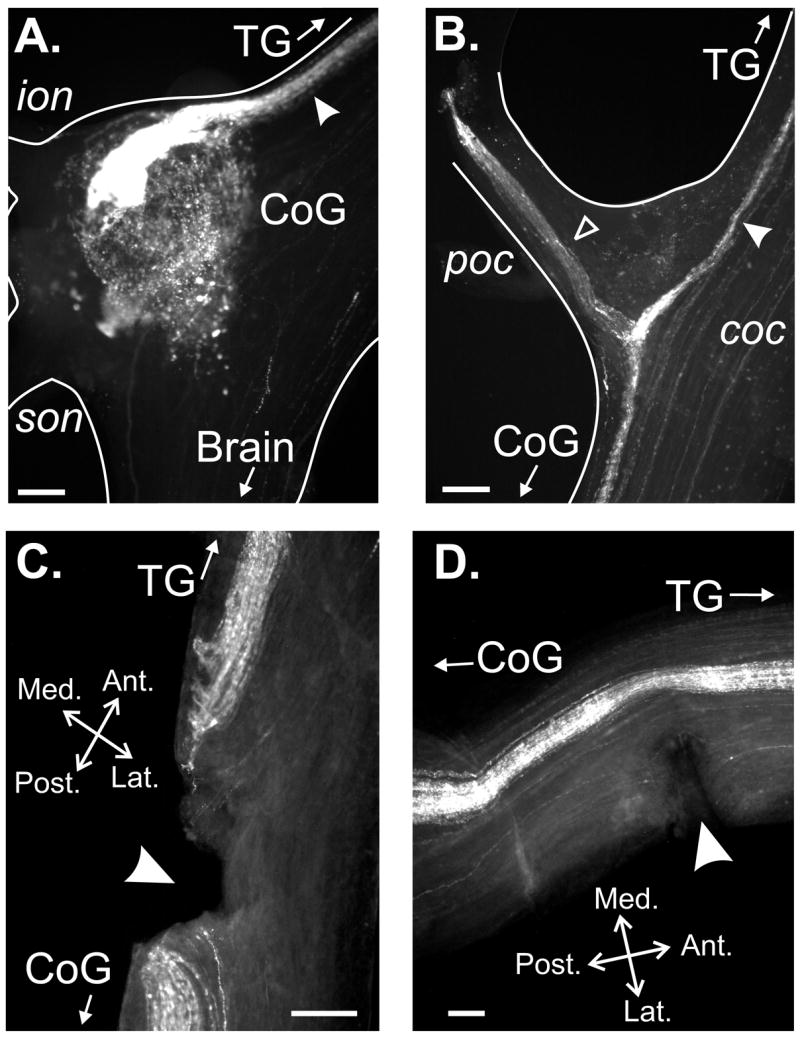
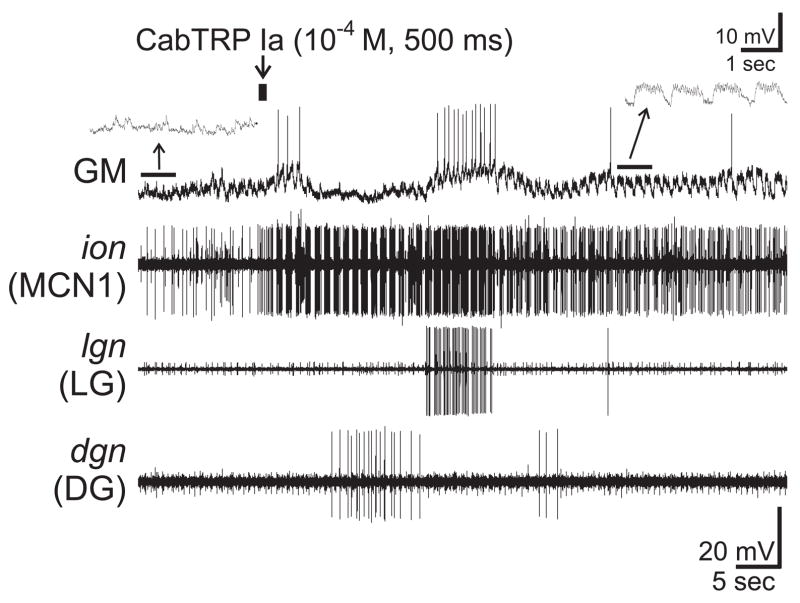
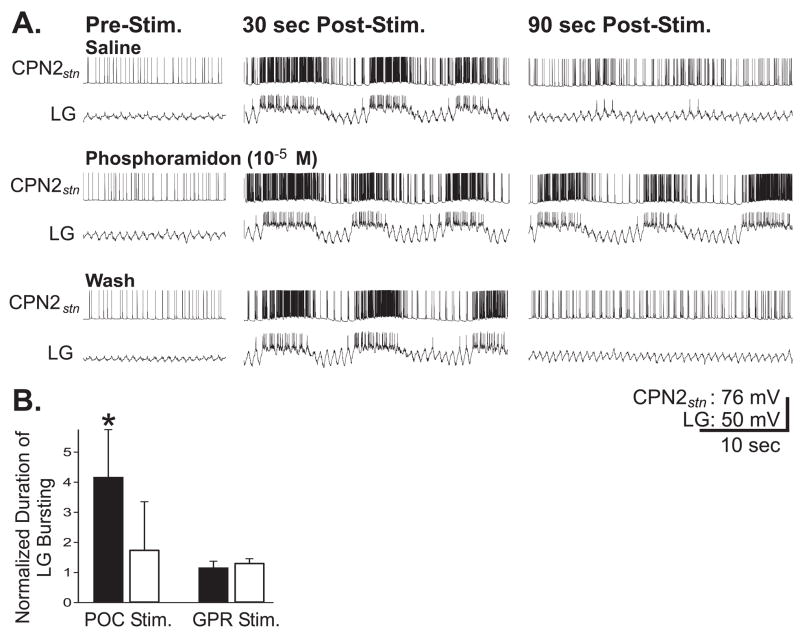
Neuronal network flexibility enables animals to respond appropriately to changes in their internal and external states. We are using the isolated crab stomatogastric nervous system to determine how extrinsic inputs contribute to network flexibility. The stomatogastric system includes the well-characterized gastric mill (chewing) and pyloric (filtering of chewed food) motor circuits in the stomatogastric ganglion. Projection neurons with somata in the commissural ganglia (CoGs) regulate these rhythms. Previous work characterized a unique gastric mill rhythm that occurred spontaneously in some preparations, but whose origin remained undetermined. This rhythm includes a distinct protractor phase activity pattern, during which a key gastric mill circuit neuron (LG neuron) and the projection neurons MCN1 and CPN2 fire in a pyloric rhythm-timed activity pattern instead of the tonic firing pattern exhibited by these neurons during previously studied gastric mill rhythms. Here we identify a new extrinsic input, the post-oesophageal commissure (POC) neurons, relatively brief stimulation (30 s) of which triggers a long-lasting (tens of minutes) activation of this novel gastric mill rhythm at least in part via its lasting activation of MCN1 and CPN2. Immunocytochemical and electrophysiological data suggest that the POC neurons excite MCN1 and CPN2 by release of the neuropeptide Cancer borealis tachykinin-related peptide Ia (CabTRP Ia). These data further suggest that the CoG arborization of the POC neurons comprises the previously identified anterior commissural organ (ACO), a CabTRP Ia-containing neurohemal organ. This endocrine organ thus appears to also have paracrine actions, including activation of a novel and lasting gastric mill rhythm.
Figures

References
-
- Alexandrowicz JS. Muscle receptor organs in the abdomen of Homarus vulgaris and Palinurus vulgaris. Quart J Micr Sci. 1951;92:163–199.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
