

Diverse pore loops of the AAA+ ClpX machine mediate unassisted and adaptor-dependent recognition of ssrA-tagged substrates
- PMID: 18313382
- PMCID: PMC2323458
- DOI: 10.1016/j.molcel.2008.02.002
Diverse pore loops of the AAA+ ClpX machine mediate unassisted and adaptor-dependent recognition of ssrA-tagged substrates
Abstract
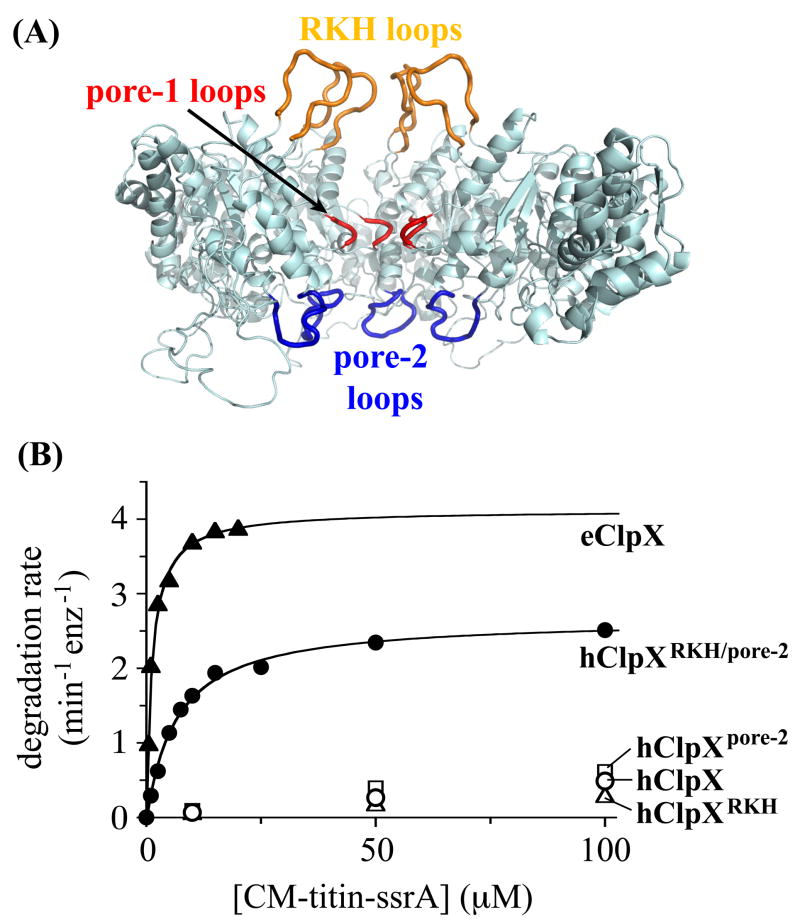
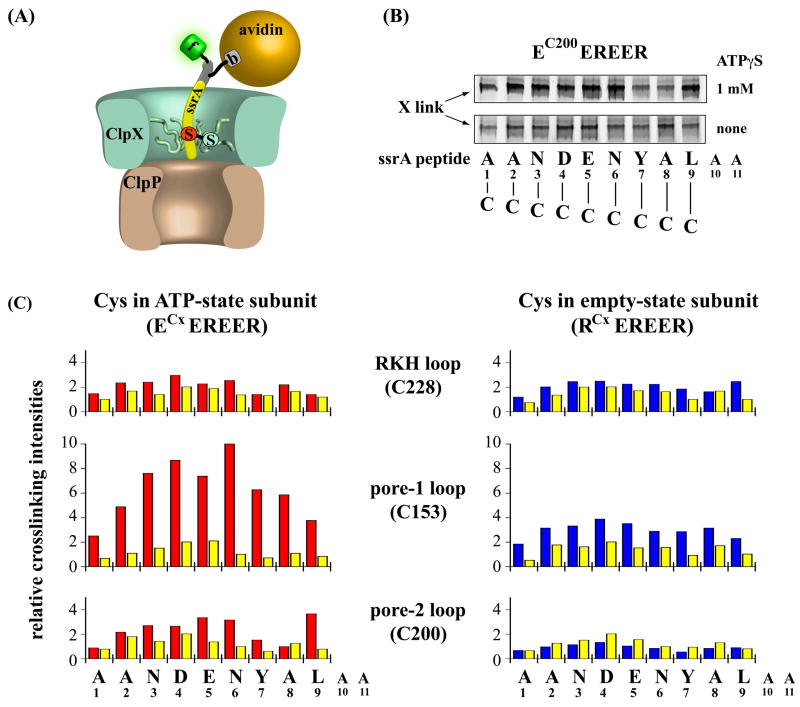
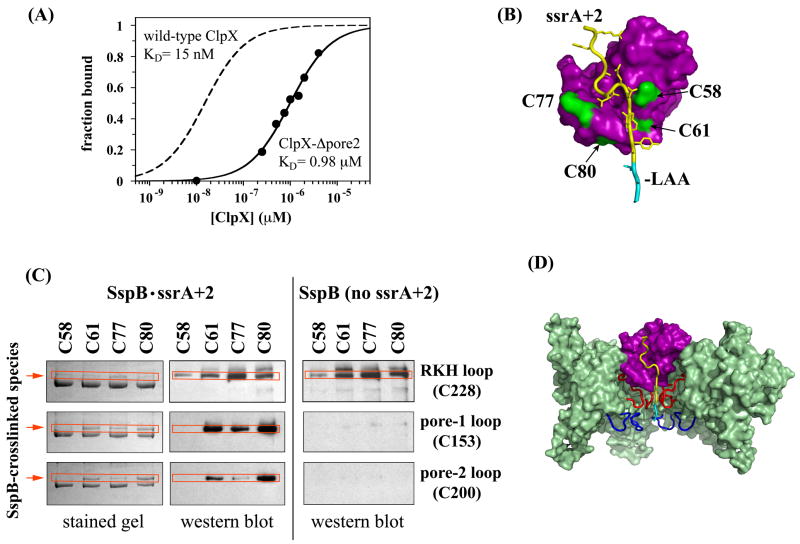
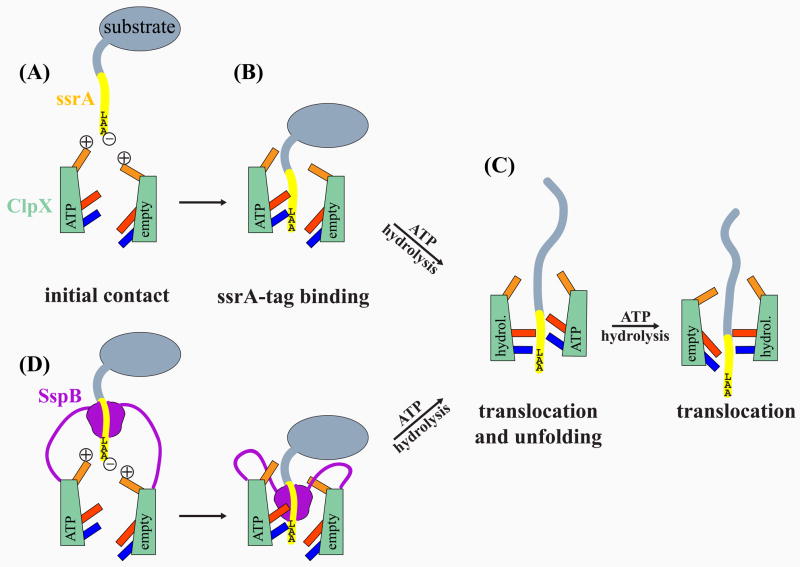
ClpX, an archetypal proteolytic AAA+ unfoldase, must engage the ssrA tags of appropriate substrates prior to ATP-dependent unfolding and translocation of the denatured polypeptide into ClpP for degradation. Here, specificity-transplant and disulfide-crosslinking experiments reveal that the ssrA tag interacts with different loops that form the top, middle, and lower portions of the central channel of the ClpX hexamer. Our results support a two-step binding mechanism, in which the top loop serves as a specificity filter and the remaining loops form a binding site for the peptide tag relatively deep within the pore. Crosslinking experiments suggest a staggered arrangement of pore loops in the hexamer and nucleotide-dependent changes in pore-loop conformations. This mechanism of initial tag binding would allow ATP-dependent conformational changes in one or more pore loops to drive peptide translocation, force unfolding, and mediate threading of the denatured protein through the ClpX pore.
Figures
References
-
- Bochtler M, Hartmann C, Song HK, Bourenkov GP, Bartunik HD, Huber R. The structures of HsIU and the ATP-dependent protease HsIU-HsIV. Nature. 2000;403:800–805. - PubMed
-
- Bolon DN, Grant RA, Baker TA, Sauer RT. Nucleotide-dependent substrate handoff from the SspB adaptor to the AAA+ ClpXP protease. Mol Cell. 2004a;16:343–50. - PubMed
-
- Bolon DN, Wah DA, Hersch GL, Baker TA, Sauer RT. Bivalent tethering of SspB to ClpXP is required for efficient substrate delivery: a protein-design study. Mol Cell. 2004b;13:443–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
