

Tocopherols modulate extraplastidic polyunsaturated fatty acid metabolism in Arabidopsis at low temperature
- PMID: 18314499
- PMCID: PMC2276453
- DOI: 10.1105/tpc.107.054718
Tocopherols modulate extraplastidic polyunsaturated fatty acid metabolism in Arabidopsis at low temperature
Abstract
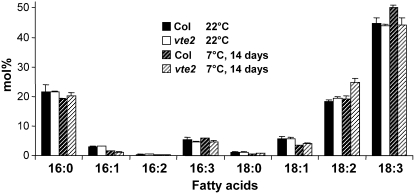
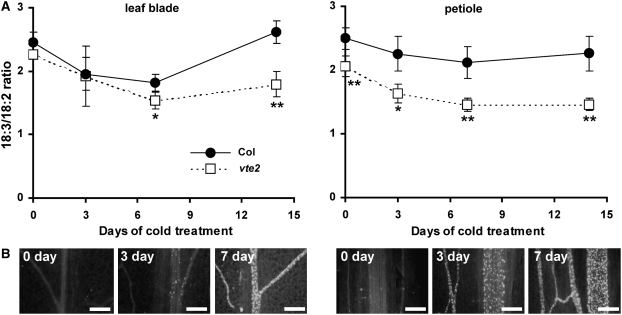
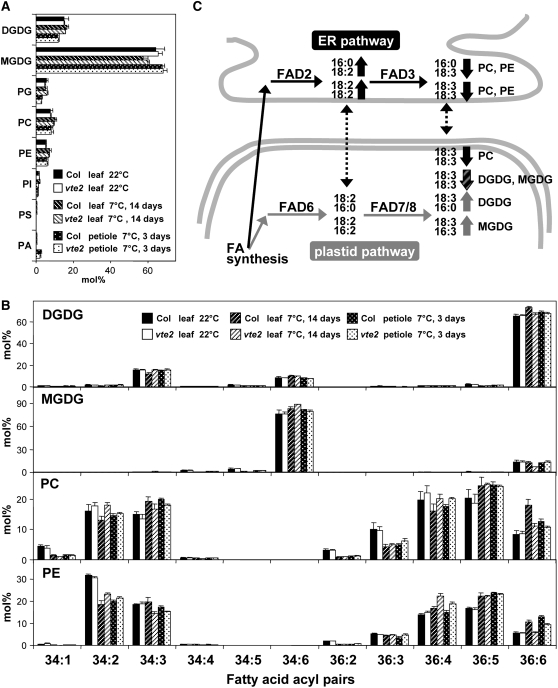
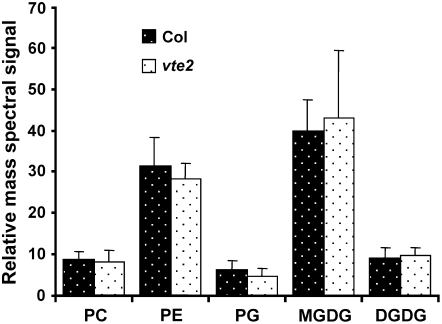
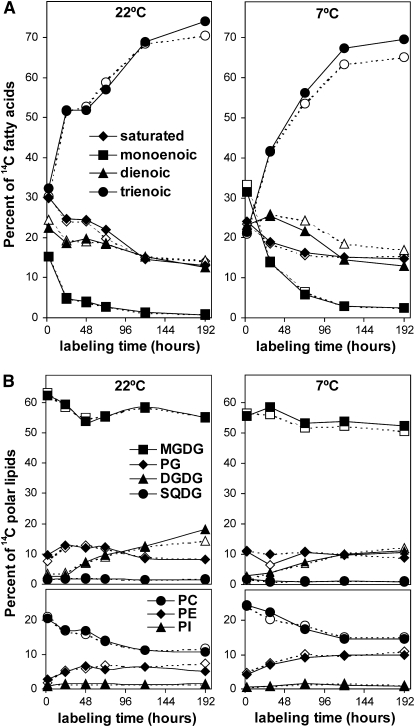
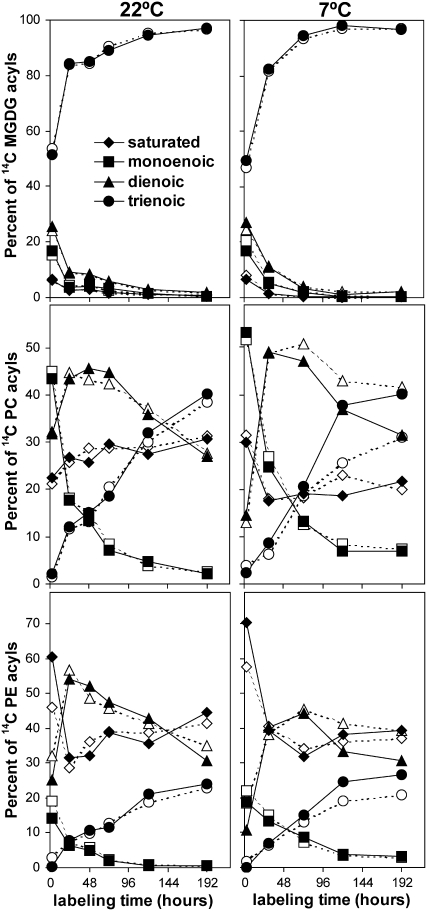
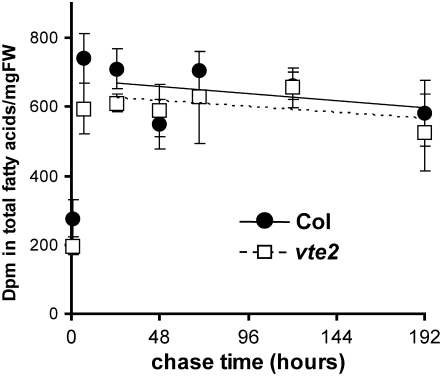
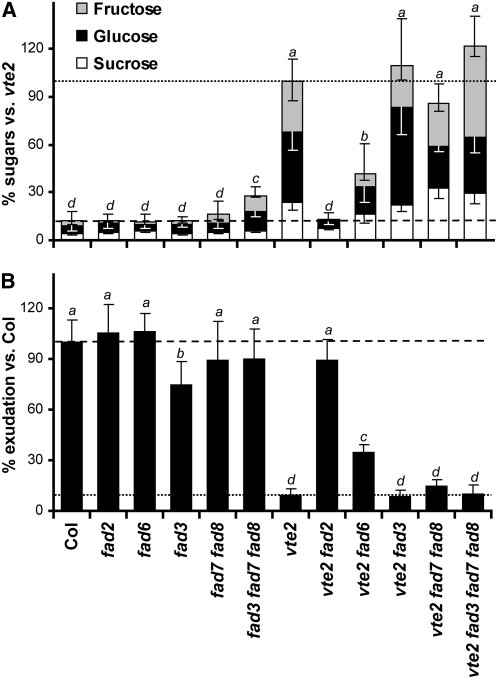
Tocopherols (vitamin E) are synthesized in plastids and have long been assumed to have essential functions restricted to these organelles. We previously reported that the vitamin e-deficient2 (vte2) mutant of Arabidopsis thaliana is defective in transfer cell wall development and photoassimilate transport at low temperature (LT). Here, we demonstrate that LT-treated vte2 has a distinct composition of polyunsaturated fatty acids (PUFAs): lower levels of linolenic acid (18:3) and higher levels of linoleic acid (18:2) compared with the wild type. Enhanced 18:3 oxidation was not involved, as indicated by the limited differences in oxidized lipid species between LT-treated vte2 and the wild type and by a lack of impact on the LT-induced vte2 phenotype in a vte2 fad3 fad7 fad8 quadruple mutant deficient in 18:3. PUFA changes in LT-treated vte2 occur primarily in phospholipids due to reduced conversion of dienoic to trienoic fatty acids in the endoplasmic reticulum (ER) pathway. Introduction of the ER fatty acid desaturase mutation, fad2, and to a lesser extent the plastidic fad6 mutation into the vte2 background suppressed the LT-induced vte2 phenotypes, including abnormal transfer cell wall development. These results provide biochemical and genetic evidence that plastid-synthesized tocopherols modulate ER PUFA metabolism early in the LT adaptation response of Arabidopsis.
Figures

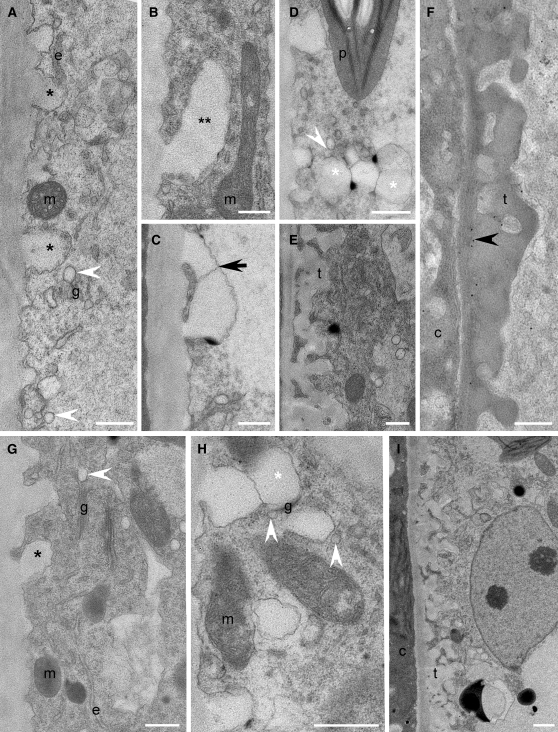
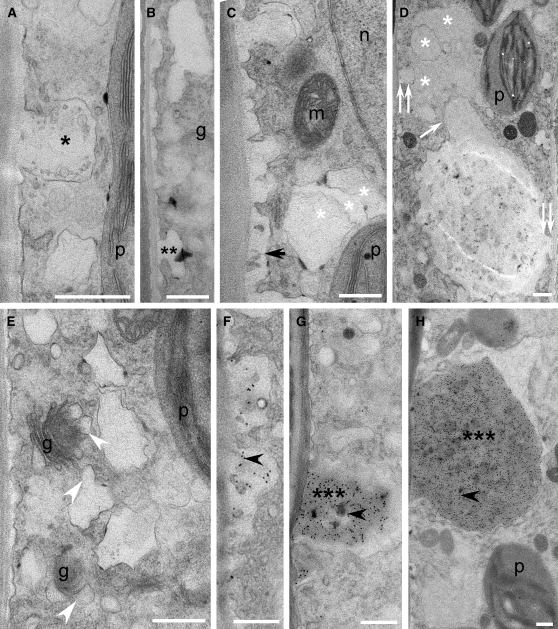
Comment in
-
Tocopherols and ER fatty acid metabolism.Plant Cell. 2008 Feb;20(2):246. doi: 10.1105/tpc.108.200212. Epub 2008 Feb 26. Plant Cell. 2008. PMID: 18314498 Free PMC article. No abstract available.
References
-
- Andersson, M.X., Goksor, M., and Sandelius, A.S. (2006). Optical manipulation reveals strong attracting forces at membrane contact sites between endoplasmic reticulum and chloroplasts. J. Biol. Chem. 282 1170–1174. - PubMed
-
- Aubert, S., Gout, E., Bligny, R., Marty-Mazars, D., Barrieu, F., Alabouvette, J., Marty, F., and Douce, R. (1996). Ultrastructural and biochemical characterization of autophagy in higher plant cells subjected to carbon deprivation: control by the supply of mitochondria with respiratory substrates. J. Cell Biol. 133 1251–1263. - PMC - PubMed
-
- Bagnall, N., Wang, X.D., Scofield, G.N., Furbank, R.T., Offler, C.E., and Patrick, J.W. (2000). Sucrose transport-related genes are expressed in both maternal and filial tissues of developing wheat grains. Aust. J. Plant Physiol. 27 1009–1020.
-
- Bonnemain, J.L., Borguin, S., Renault, S., Offler, C.E., and Fincher, G.B. (1991). Transfer cells: Structure and physiology. In Recent Advances in Phloem Transport and Assimilate Compartmentation, L. Bonnemain and S. Delrot, eds (Paris: Presses Académiques), pp. 74–83.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
