

Role of SLV in SLI substrate recognition by the Neurospora VS ribozyme
- PMID: 18314503
- PMCID: PMC2271362
- DOI: 10.1261/rna.824308
Role of SLV in SLI substrate recognition by the Neurospora VS ribozyme
Abstract
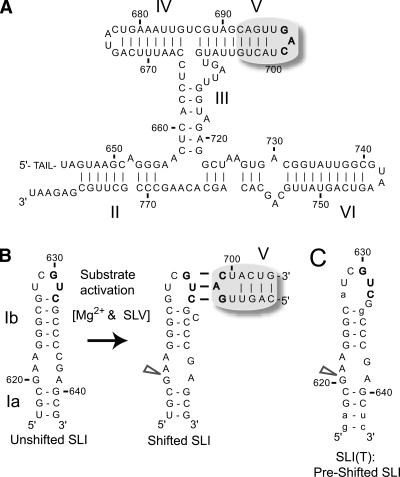
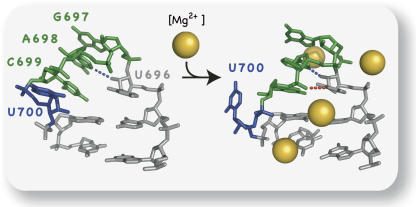
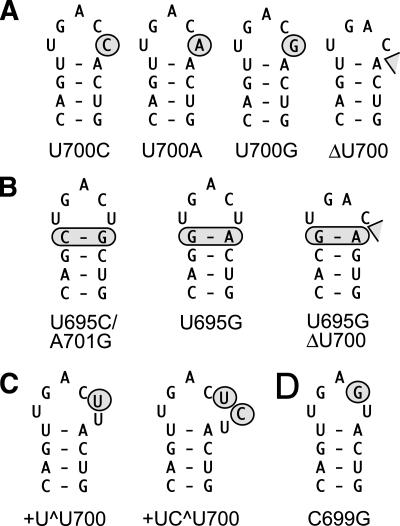
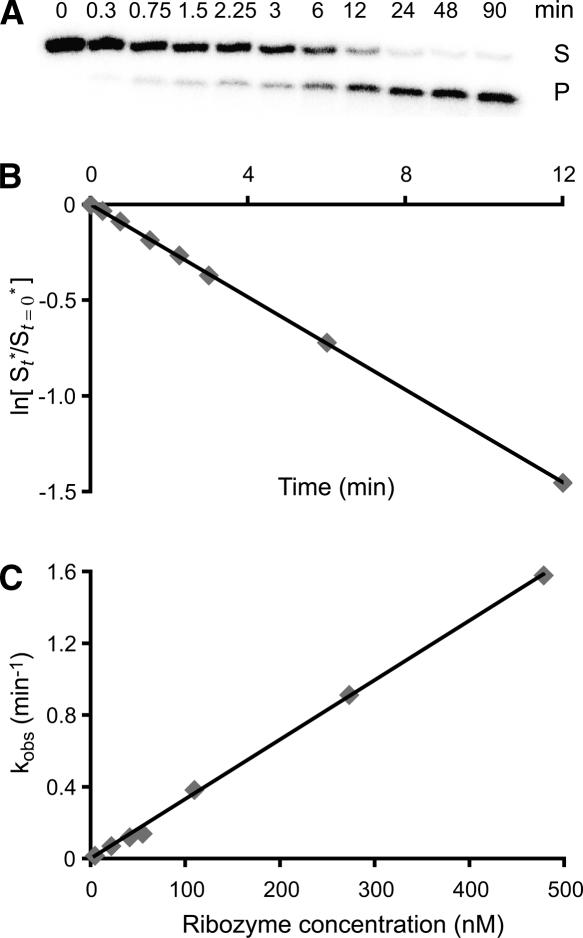
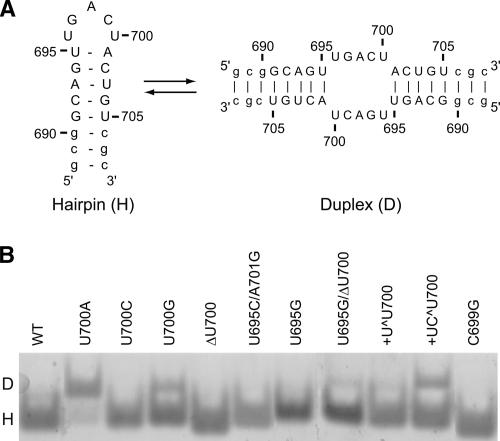
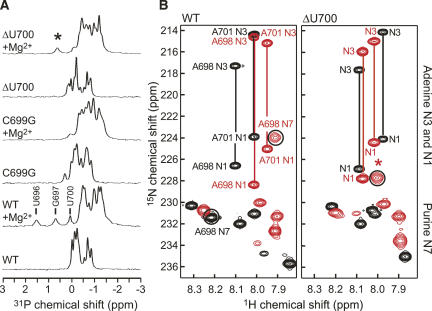
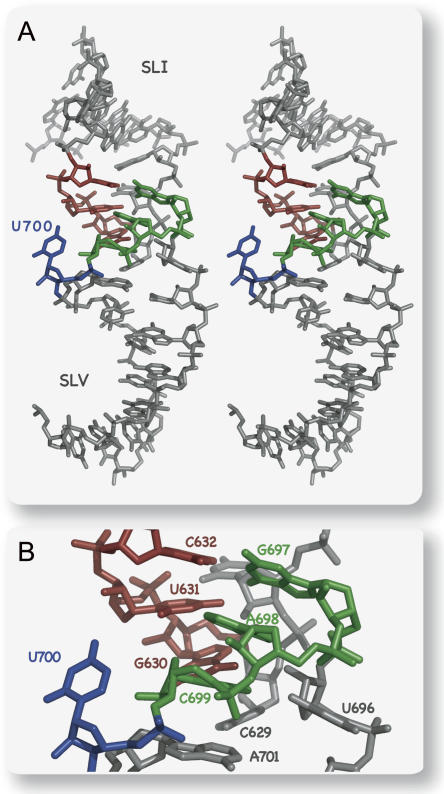
Substrate recognition by the VS ribozyme involves a magnesium-dependent loop/loop interaction between the SLI substrate and the SLV hairpin from the catalytic domain. Recent NMR studies of SLV demonstrated that magnesium ions stabilize a U-turn loop structure and trigger a conformational change for the extruded loop residue U700, suggesting a role for U700 in SLI recognition. Here, we kinetically characterized VS ribozyme mutants to evaluate the contribution of U700 and other SLV loop residues to SLI recognition. To help interpret the kinetic data, we structurally characterized the SLV mutants by NMR spectroscopy and generated a three-dimensional model of the SLI/SLV complex by homology modeling with MC-Sym. We demonstrated that the mutation of U700 by A, C, or G does not significantly affect ribozyme activity, whereas deletion of U700 dramatically impairs this activity. The U700 backbone is likely important for SLI recognition, but does not appear to be required for either the structural integrity of the SLV loop or for direct interactions with SLI. Thus, deletion of U700 may affect other aspects of SLI recognition, such as magnesium ion binding and SLV loop dynamics. As part of our NMR studies, we developed a convenient assay based on detection of unusual (31)P and (15)N N7 chemical shifts to probe the formation of U-turn structures in RNAs. Our model of the SLI/SLV complex, which is compatible with biochemical data, leads us to propose novel interactions at the loop I/loop V interface.
Figures
References
-
- Andersen, A., Collins, R.A. Rearrangement of a stable RNA secondary structure during VS ribozyme catalysis. Mol. Cell. 2000;5:469–478. - PubMed
-
- Beattie, T.L., Collins, R.A. Identification of functional domains in the self-cleaving Neurospora VS ribozyme using damage selection. J. Mol. Biol. 1997;267:830–840. - PubMed
-
- Cabello-Villegas, J., Tworowska, I., Nikonowicz, E.P. Metal ion stabilization of the U-turn of the A37 N6-dimethylallyl-modified anticodon stem–loop of Escherichia coli tRNAPhe . Biochemistry. 2004;43:55–66. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources