

Cdk11 is a RanGTP-dependent microtubule stabilization factor that regulates spindle assembly rate
- PMID: 18316407
- PMCID: PMC2265385
- DOI: 10.1083/jcb.200706189
Cdk11 is a RanGTP-dependent microtubule stabilization factor that regulates spindle assembly rate
Abstract
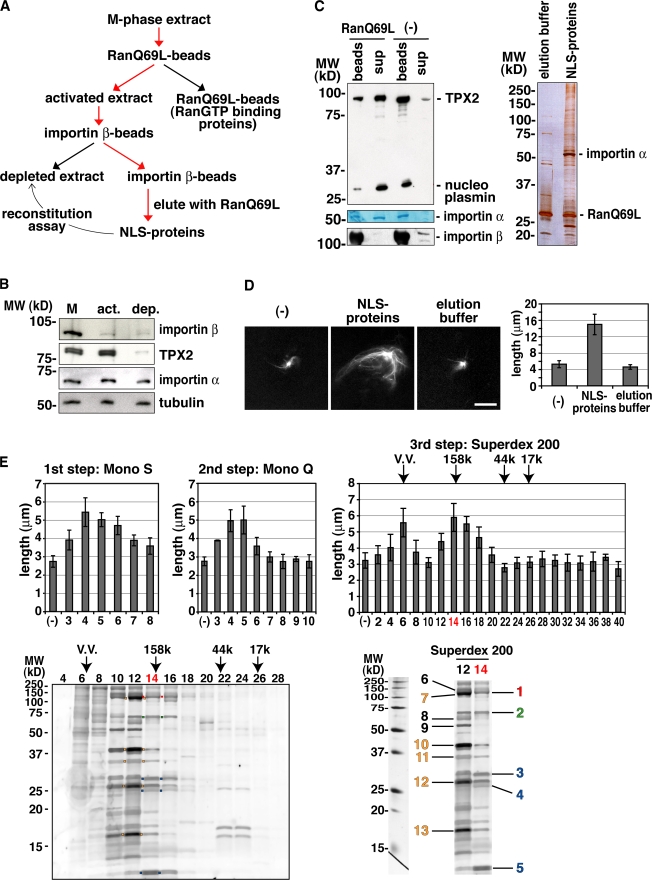
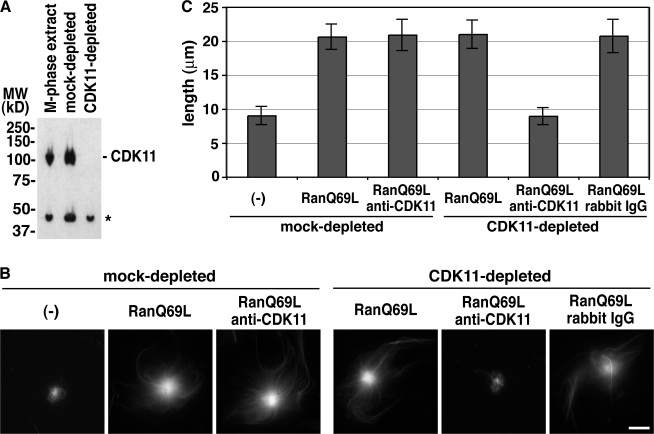
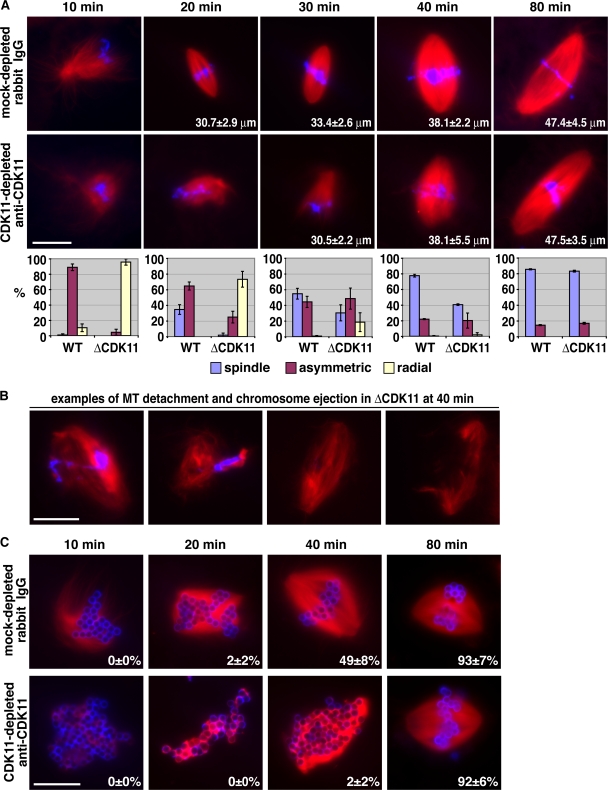
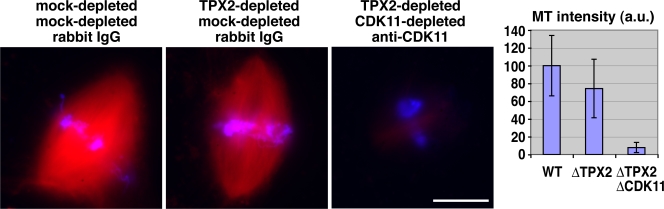
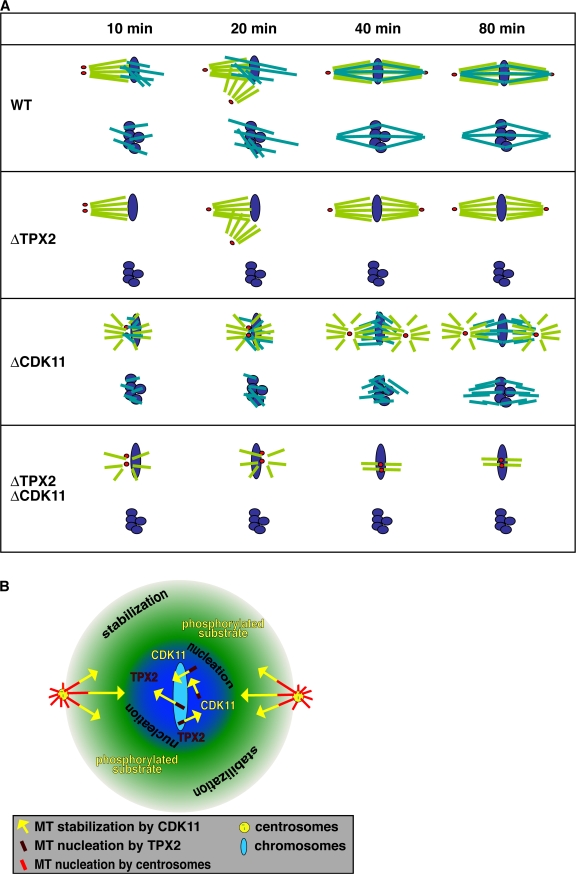
Production of Ran-guanosine triphosphate (GTP) around chromosomes induces local nucleation and plus end stabilization of microtubules (MTs). The nuclear protein TPX2 is required for RanGTP-dependent MT nucleation. To find the MT stabilizer, we affinity purify nuclear localization signal (NLS)-containing proteins from Xenopus laevis egg extracts. This NLS protein fraction contains the MT stabilization activity. After further purification, we used mass spectrometry to identify proteins in active fractions, including cyclin-dependent kinase 11 (Cdk11). Cdk11 localizes on spindle poles and MTs in Xenopus culture cells and egg extracts. Recombinant Cdk11 demonstrates RanGTP-dependent MT stabilization activity, whereas a kinase-dead mutant does not. Inactivation of Cdk11 in egg extracts blocks RanGTP-dependent MT stabilization and dramatically decreases the spindle assembly rate. Simultaneous depletion of TPX2 completely inhibits centrosome-dependent spindle assembly. Our results indicate that Cdk11 is responsible for RanGTP-dependent MT stabilization around chromosomes and that this local stabilization is essential for normal rates of spindle assembly and spindle function.
Figures
References
-
- Bastiaens, P., M. Caudron, P. Niethammer, and E. Karsenti. 2006. Gradients in the self-organization of the mitotic spindle. Trends Cell Biol. 16:125–134. - PubMed
-
- Berke, J.D., V. Sgambato, P.P. Zhu, B. Lavoie, M. Vincent, M. Krause, and S.E. Hyman. 2001. Dopamine and glutamate induce distinct striatal splice forms of Ania-6, an RNA polymerase II-associated cyclin. Neuron. 32:277–287. - PubMed
-
- Burbank, K.S., T.J. Mitchison, and D.S. Fisher. 2007. Slide-and-cluster models for spindle assembly. Curr. Biol. 17:1373–1383. - PubMed
-
- Carazo-Salas, R.E., G. Guarguaglini, O.J. Gruss, A. Segref, E. Karsenti, and I.W. Mattaj. 1999. Generation of GTP-bound Ran by RCC1 is required for chromatin-induced mitotic spindle formation. Nature. 400:178–181. - PubMed
-
- Carazo-Salas, R.E., O.J. Gruss, I.W. Mattaj, and E. Karsenti. 2001. Ran-GTP coordinates regulation of microtubule nucleation and dynamics during mitotic-spindle assembly. Nat. Cell Biol. 3:228–234. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
