

Methylation of RUNX1 by PRMT1 abrogates SIN3A binding and potentiates its transcriptional activity
- PMID: 18316480
- PMCID: PMC2259033
- DOI: 10.1101/gad.1632608
Methylation of RUNX1 by PRMT1 abrogates SIN3A binding and potentiates its transcriptional activity
Abstract
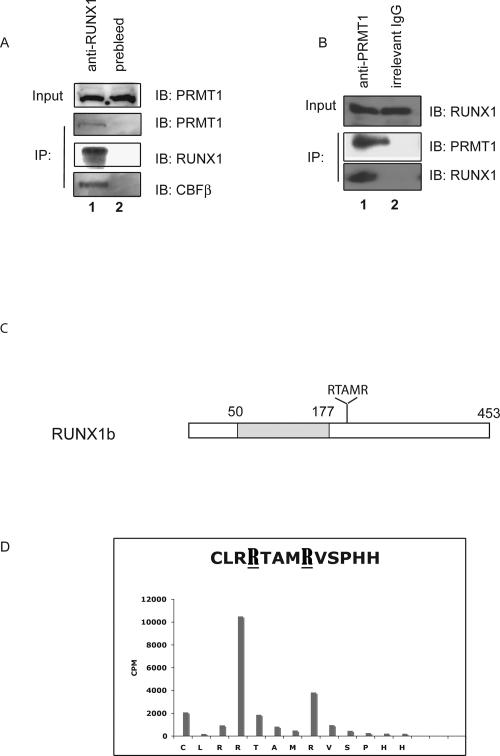
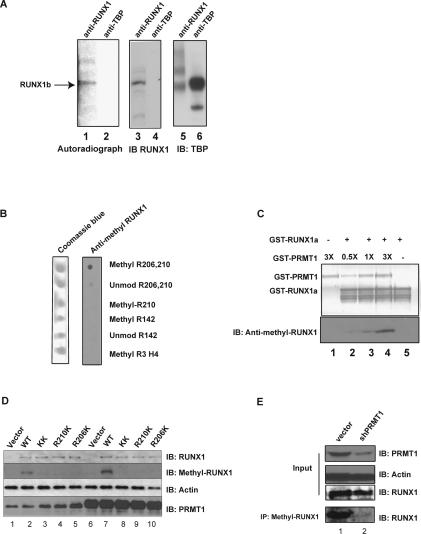
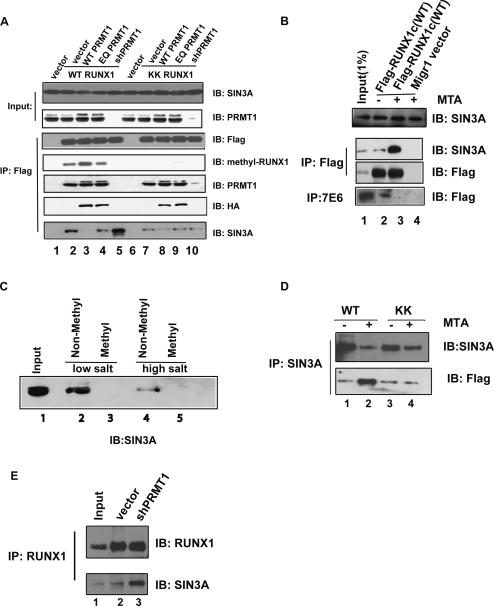
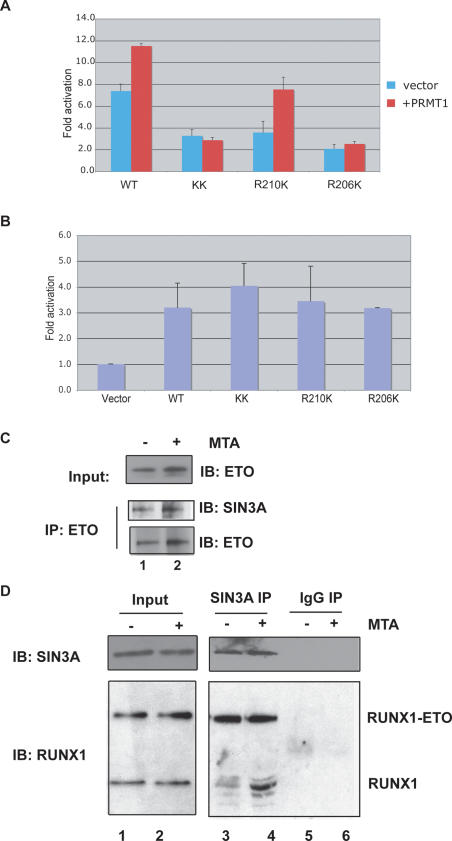
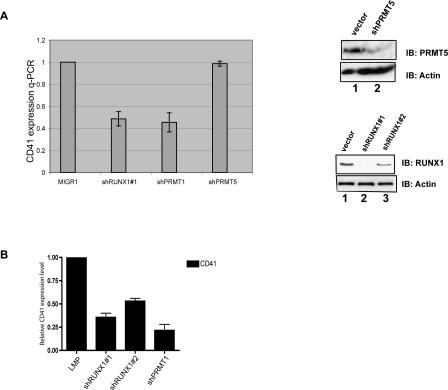
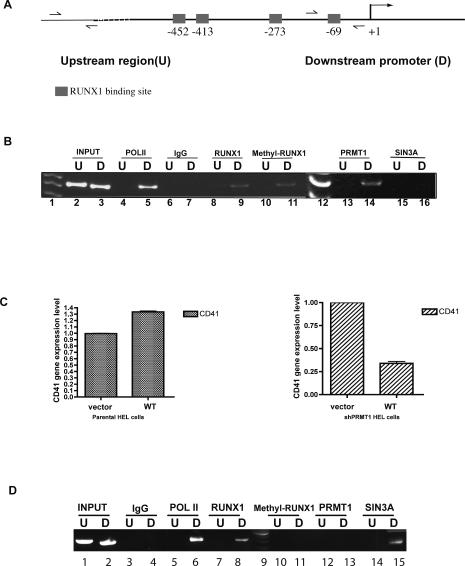
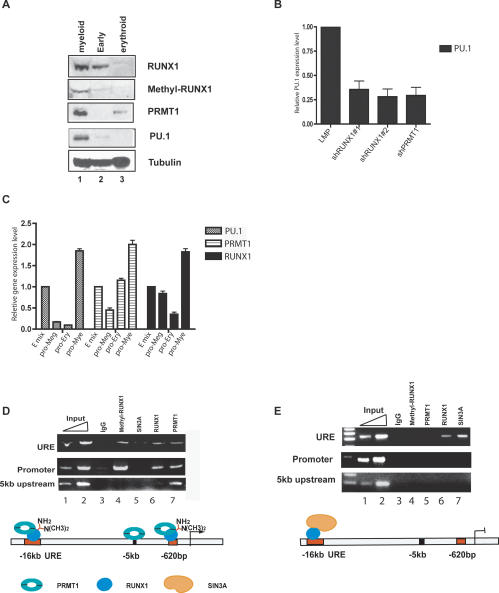
RUNX1/AML1 is required for the development of definitive hematopoiesis, and its activity is altered by mutations, deletions, and chromosome translocations in human acute leukemia. RUNX1 function can be regulated by post-translational modifications and protein-protein interactions. We show that RUNX1 is arginine-methylated in vivo by the arginine methyltransferase PRMT1, and that PRMT1 serves as a transcriptional coactivator for RUNX1 function. Using mass spectrometry, and a methyl-arginine-specific antibody, we identified two arginine residues (R206 and R210) within the region of RUNX1 that interact with the corepressor SIN3A and are methylated by PRMT1. PRMT1- dependent methylation of RUNX1 at these arginine residues abrogates its association with SIN3A, whereas shRNA against PRMT1 (or use of a methyltransferase inhibitor) enhances this association. We find arginine-methylated RUNX1 on the promoters of two bona fide RUNX1 target genes, CD41 and PU.1 and show that shRNA against PRMT1 or RUNX1 down-regulates their expression. These arginine methylation sites and the dynamic regulation of corepressor binding are lost in the leukemia-associated RUNX1-ETO fusion protein, which likely contributes to its dominant inhibitory activity.
Figures
References
-
- An W., Kim J., Roeder R.G. Ordered cooperative functions of PRMT1, p300, and CARM1 in transcriptional activation by p53. Cell. 2004;117:735–748. - PubMed
-
- Bedford M.T., Richard S. Arginine methylation an emerging regulator of protein function. Mol. Cell. 2005;18:263–272. - PubMed
-
- Boisvert F.M., Cote J., Boulanger M.C., Richard S. A proteomic analysis of arginine-methylated protein complexes. Mol. Cell. Proteomics. 2003;2:1319–1330. - PubMed
-
- Bruhn L., Munnerlyn A., Grosschedl R. ALY, a context-dependent coactivator of LEF-1 and AML-1, is required for TCRα enhancer function. Genes & Dev. 1997;11:640–653. - PubMed
-
- Chakraborty S., Sinha K.K., Senyuk V., Nucifora G. SUV39H1 interacts with AML1 and abrogates AML1 transactivity. AML1 is methylated in vivo. Oncogene. 2003;22:5229–5237. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases