

HMGA1 is a molecular determinant of chemoresistance to gemcitabine in pancreatic adenocarcinoma
- PMID: 18316571
- PMCID: PMC2652398
- DOI: 10.1158/1078-0432.CCR-07-1450
HMGA1 is a molecular determinant of chemoresistance to gemcitabine in pancreatic adenocarcinoma
Erratum in
-
Editor's Note: HMGA1 Is a Molecular Determinant of Chemoresistance to Gemcitabine in Pancreatic Adenocarcinoma.Clin Cancer Res. 2025 May 15;31(10):2064. doi: 10.1158/1078-0432.CCR-25-0819. Clin Cancer Res. 2025. PMID: 40370047 No abstract available.
Abstract
Purpose: HMGA1 proteins are architectural transcription factors that are overexpressed by pancreatic adenocarcinomas. We previously have shown that RNA interference targeting the HMGA1 gene may represent a potential chemosensitizing strategy in pancreatic adenocarcinoma cells. In this study, we tested the hypothesis that HMGA1 promotes chemoresistance to gemcitabine in pancreatic cancer cells.
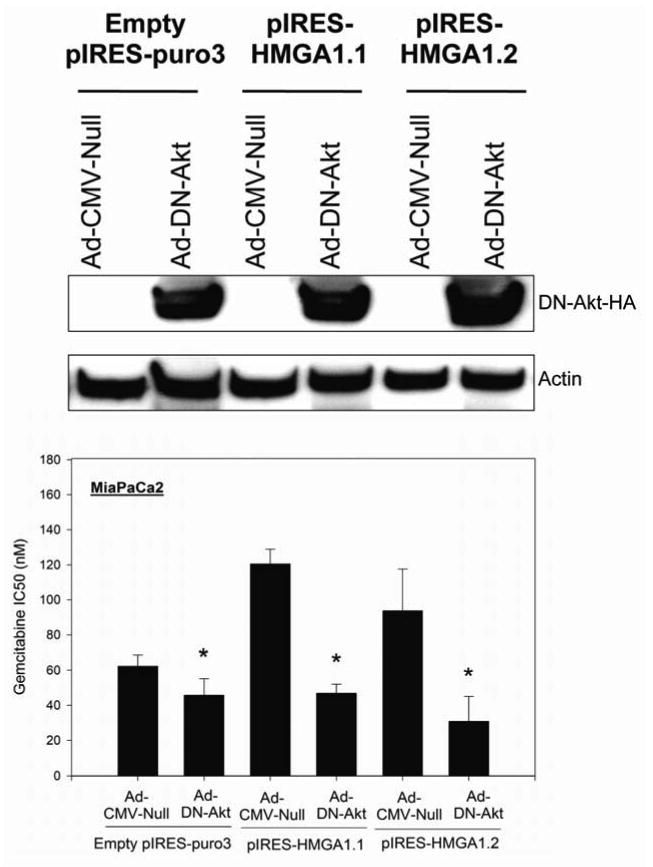
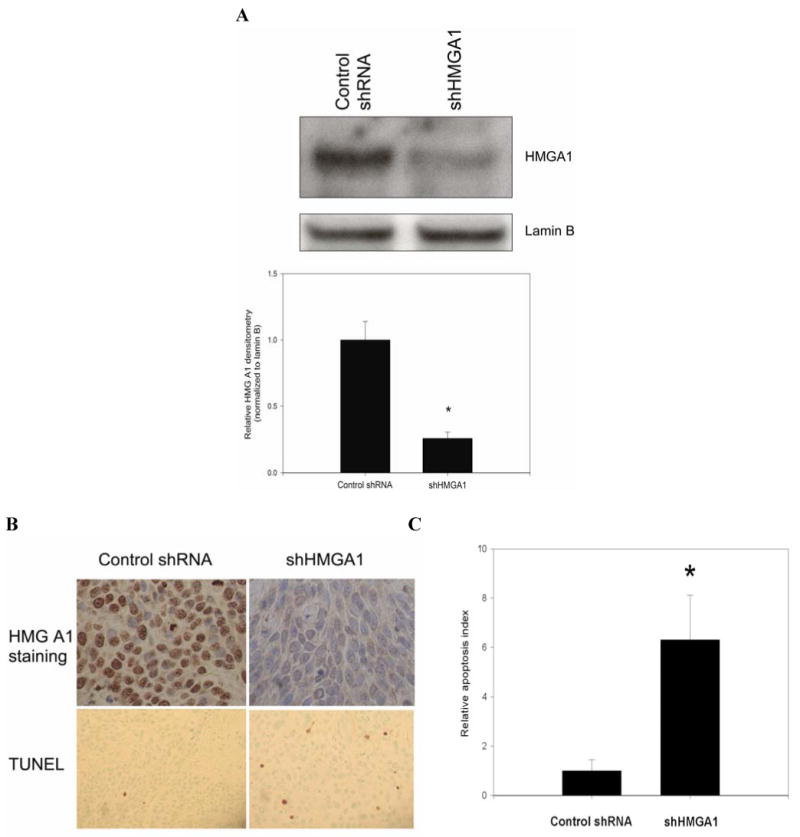
Experimental design and results: Stable short hairpin RNA-mediated HMGA1 silencing in BxPC3 and MiaPaCa2 cells promoted chemosensitivity to gemcitabine, with reductions in gemcitabine IC(50) and increases in gemcitabine-induced apoptosis and caspase-3 activation. In contrast, forced HMGA1 overexpression in MiaPaCa2 cells promoted chemoresistance to gemcitabine, with increases in gemcitabine IC(50) and reductions in gemcitabine-induced apoptosis and caspase-3 activation. Dominant negative Akt abrogated HMGA1 overexpression-induced increases in chemoresistance to gemcitabine. Finally, HMGA1 silencing promoted chemosensitivity to gemcitabine in vivo in a nude mouse xenograft model of pancreatic adenocarcinoma.
Conclusion: Our findings suggest that HMGA1 promotes chemoresistance to gemcitabine through an Akt-dependent mechanism. Targeted therapies directed at HMGA1 represent a potential strategy for ameliorating chemoresistance in pancreatic adenocarcinoma.
Figures




Comment in
-
Gemcitabine resistance in pancreatic cancer: picking the key players.Clin Cancer Res. 2008 Mar 1;14(5):1284-5. doi: 10.1158/1078-0432.CCR-07-2247. Clin Cancer Res. 2008. PMID: 18316544 No abstract available.
References
-
- Jemal A, Siegel R, Ward E. Cancer statistics, 2006. CA Cancer J Clin. 2006;56:106–30. - PubMed
-
- Sener SF, Fremgen A, Menck HR, Winchester DP. Pancreatic cancer: a report of treatment and survival trends for 100,313 patients diagnosed from 1985-1995, using the National Cancer Database. J Am Coll Surg. 1999;189:1–7. - PubMed
-
- Thanos D, Maniatis T. Virus induction of human IFN beta gene expression requires the assembly of an enhanceosome. Cell. 1995;83:1091–100. - PubMed
-
- Du W, Thanos D, Maniatis T. Mechanisms of transcriptional synergism between distinct virus-inducible enhancer elements. Cell. 1993;74:887–98. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
