

A novel methyltransferase methylates Cucumber mosaic virus 1a protein and promotes systemic spread
- PMID: 18321966
- PMCID: PMC2346749
- DOI: 10.1128/JVI.02518-07
A novel methyltransferase methylates Cucumber mosaic virus 1a protein and promotes systemic spread
Abstract
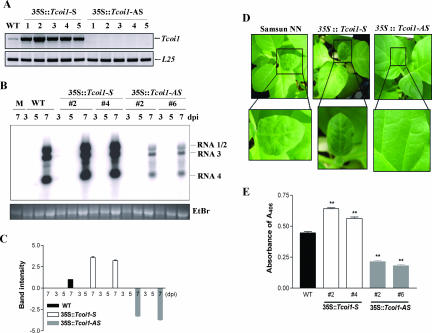
In mammalian and yeast systems, methyltransferases have been implicated in the regulation of diverse processes, such as protein-protein interactions, protein localization, signal transduction, RNA processing, and transcription. The Cucumber mosaic virus (CMV) 1a protein is essential not only for virus replication but also for movement. Using a yeast two-hybrid system with tobacco plants, we have identified a novel gene encoding a methyltransferase that interacts with the CMV 1a protein and have designated this gene Tcoi1 (tobacco CMV 1a-interacting protein 1). Tcoi1 specifically interacted with the methyltransferase domain of CMV 1a, and the expression of Tcoi1 was increased by CMV inoculation. Biochemical studies revealed that the interaction of Tcoi1 with CMV 1a protein was direct and that Tcoi1 methylated CMV 1a protein both in vitro and in vivo. The CMV 1a binding activity of Tcoi1 is in the C-terminal domain, which shows the methyltransferase activity. The overexpression of Tcoi1 enhanced the CMV infection, while the reduced expression of Tcoi1 decreased virus infectivity. These results suggest that Tcoi1 controls the propagation of CMV through an interaction with the CMV 1a protein.
Figures

Similar articles
-
A zinc finger protein Tsip1 controls Cucumber mosaic virus infection by interacting with the replication complex on vacuolar membranes of the tobacco plant.New Phytol. 2011 Aug;191(3):746-762. doi: 10.1111/j.1469-8137.2011.03717.x. Epub 2011 Apr 8. New Phytol. 2011. PMID: 21477206
-
Novel protein kinase interacts with the Cucumber mosaic virus 1a methyltransferase domain.Biochem Biophys Res Commun. 2006 Feb 3;340(1):228-35. doi: 10.1016/j.bbrc.2005.11.178. Epub 2005 Dec 9. Biochem Biophys Res Commun. 2006. PMID: 16360640
-
Isolation of a putative tobacco host factor interacting with cucumber mosaic virus-encoded 2b protein by yeast two-hybrid screening.Mol Cells. 1999 Oct 31;9(5):548-55. Mol Cells. 1999. PMID: 10597045
-
Arabidopsis tonoplast proteins TIP1 and TIP2 interact with the cucumber mosaic virus 1a replication protein.J Gen Virol. 2006 Nov;87(Pt 11):3425-3431. doi: 10.1099/vir.0.82252-0. J Gen Virol. 2006. PMID: 17030879
-
Replication of cucumber mosaic virus RNA 1 in cis requires functional helicase-like motifs of the 1a protein.Virus Res. 2011 Jun;158(1-2):271-6. doi: 10.1016/j.virusres.2011.03.008. Epub 2011 Mar 23. Virus Res. 2011. PMID: 21402113
Cited by
-
The NIa-Protease Protein Encoded by the Pepper Mottle Virus Is a Pathogenicity Determinant and Releases DNA Methylation of Nicotiana benthamiana.Front Microbiol. 2020 Feb 21;11:102. doi: 10.3389/fmicb.2020.00102. eCollection 2020. Front Microbiol. 2020. PMID: 32153517 Free PMC article.
-
Four sequence positions of the movement protein of Cucumber mosaic virus determine the virulence against cmv1-mediated resistance in melon.Mol Plant Pathol. 2015 Sep;16(7):675-84. doi: 10.1111/mpp.12225. Epub 2015 Jan 29. Mol Plant Pathol. 2015. PMID: 25470079 Free PMC article.
-
Arabidopsis Pumilio protein APUM5 suppresses Cucumber mosaic virus infection via direct binding of viral RNAs.Proc Natl Acad Sci U S A. 2013 Jan 8;110(2):779-84. doi: 10.1073/pnas.1214287110. Epub 2012 Dec 26. Proc Natl Acad Sci U S A. 2013. PMID: 23269841 Free PMC article.
-
Cucumber Mosaic Virus 1a Protein Interacts with the Tobacco SHE1 Transcription Factor and Partitions between the Nucleus and the Tonoplast Membrane.Plant Pathol J. 2021 Apr;37(2):182-193. doi: 10.5423/PPJ.FT.03.2021.0045. Epub 2021 Apr 1. Plant Pathol J. 2021. PMID: 33866760 Free PMC article.
-
Analysis of two strains of Peanut stunt virus: satRNA-associated and satRNA free.Virus Genes. 2012 Jun;44(3):513-21. doi: 10.1007/s11262-012-0729-6. Epub 2012 Mar 4. Virus Genes. 2012. PMID: 22392626
References
-
- Aletta, J. M., T. R. Cimato, and M. J. Ettinger. 1998. Protein methylation: a signal event in post-translational modification. Trends Biochem. Sci. 2389-91. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl. 1995. Short protocols in molecular biology. Wiley, New York, NY.
-
- Baba, S. W., G. I. Belogrudov, J. C. Lee, P. T. Lee, J. Strahan, J. N. Shepherd, and C. F. Clarke. 2004. Yeast Coq5 C-methyltransferase is required for stability of other polypeptides involved in coenzyme Q biosynthesis. J. Biol. Chem. 27910052-10059. - PubMed
-
- Borchardt, R. T., B. T. Keller, and U. Patel-Thombre. 1984. Neplanocin A. A potent inhibitor of S-adenosylhomocysteine hydrolase and of vaccinia virus multiplication in mouse L929 cells. J. Biol. Chem. 2594353-4358. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
