

A fusion-intermediate state of HIV-1 gp41 targeted by broadly neutralizing antibodies
- PMID: 18322015
- PMCID: PMC2268799
- DOI: 10.1073/pnas.0800255105
A fusion-intermediate state of HIV-1 gp41 targeted by broadly neutralizing antibodies
Abstract
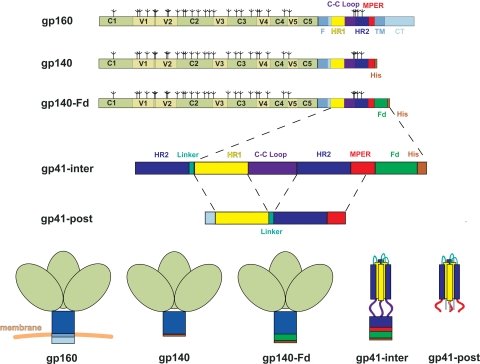
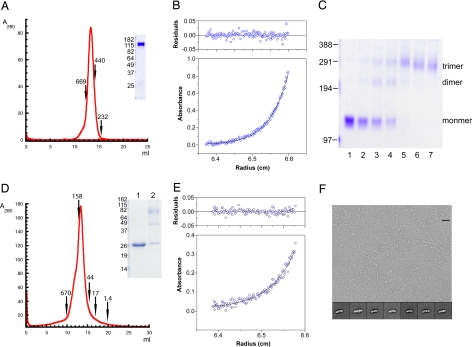
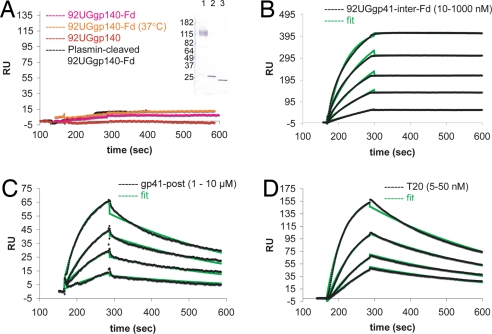
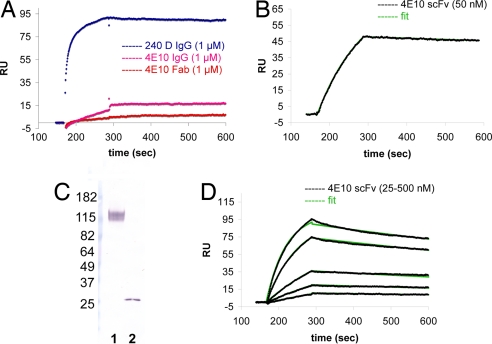
Most antibodies induced by HIV-1 are ineffective at preventing initiation or spread of infection because they are either nonneutralizing or narrowly isolate-specific. Rare, "broadly neutralizing" antibodies have been detected that recognize relatively conserved regions on the envelope glycoprotein. Using stringently characterized, homogeneous preparations of trimeric HIV-1 envelope protein in relevant conformations, we have analyzed the molecular mechanism of neutralization by two of these antibodies, 2F5 and 4E10. We find that their epitopes, in the membrane-proximal segment of the envelope protein ectodomain, are exposed only on a form designed to mimic an intermediate state during viral entry. These results help explain the rarity of 2F5- and 4E10-like antibody responses and suggest a strategy for eliciting them.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
HIV type 1 Env precursor cleavage state affects recognition by both neutralizing and nonneutralizing gp41 antibodies.AIDS Res Hum Retroviruses. 2011 Aug;27(8):877-87. doi: 10.1089/AID.2010.0281. Epub 2011 Jan 19. AIDS Res Hum Retroviruses. 2011. PMID: 21158699 Free PMC article.
-
Direct antibody access to the HIV-1 membrane-proximal external region positively correlates with neutralization sensitivity.J Virol. 2011 Aug;85(16):8217-26. doi: 10.1128/JVI.00756-11. Epub 2011 Jun 8. J Virol. 2011. PMID: 21653673 Free PMC article.
-
Analysis of the human immunodeficiency virus type 1 gp41 membrane proximal external region arrayed on hepatitis B surface antigen particles.Virology. 2008 Mar 30;373(1):72-84. doi: 10.1016/j.virol.2007.11.005. Epub 2007 Dec 26. Virology. 2008. PMID: 18155743 Free PMC article.
-
Progress towards the development of a HIV-1 gp41-directed vaccine.Curr HIV Res. 2004 Apr;2(2):193-204. doi: 10.2174/1570162043484933. Curr HIV Res. 2004. PMID: 15078183 Review.
-
The membrane-proximal external region of the human immunodeficiency virus type 1 envelope: dominant site of antibody neutralization and target for vaccine design.Microbiol Mol Biol Rev. 2008 Mar;72(1):54-84, table of contents. doi: 10.1128/MMBR.00020-07. Microbiol Mol Biol Rev. 2008. PMID: 18322034 Free PMC article. Review.
Cited by
-
Isotype modulates epitope specificity, affinity, and antiviral activities of anti-HIV-1 human broadly neutralizing 2F5 antibody.Proc Natl Acad Sci U S A. 2012 Jul 31;109(31):12680-5. doi: 10.1073/pnas.1200024109. Epub 2012 Jun 20. Proc Natl Acad Sci U S A. 2012. PMID: 22723360 Free PMC article.
-
Induction of HIV-1 broad neutralizing antibodies in 2F5 knock-in mice: selection against membrane proximal external region-associated autoreactivity limits T-dependent responses.J Immunol. 2013 Sep 1;191(5):2538-50. doi: 10.4049/jimmunol.1300971. Epub 2013 Aug 5. J Immunol. 2013. PMID: 23918977 Free PMC article.
-
Regulation of epitope exposure in the gp41 membrane-proximal external region through interactions at the apex of HIV-1 Env.PLoS Pathog. 2022 May 18;18(5):e1010531. doi: 10.1371/journal.ppat.1010531. eCollection 2022 May. PLoS Pathog. 2022. PMID: 35584191 Free PMC article.
-
Examination of the contributions of size and avidity to the neutralization mechanisms of the anti-HIV antibodies b12 and 4E10.Proc Natl Acad Sci U S A. 2009 May 5;106(18):7385-90. doi: 10.1073/pnas.0811427106. Epub 2009 Apr 16. Proc Natl Acad Sci U S A. 2009. PMID: 19372381 Free PMC article.
-
Mechanistic studies of a T20-dependent human immunodeficiency virus type 1 variant.J Virol. 2008 Aug;82(15):7735-40. doi: 10.1128/JVI.02524-07. Epub 2008 May 14. J Virol. 2008. PMID: 18480456 Free PMC article.
References
-
- Wei X, et al. Antibody neutralization and escape by HIV-1. Nature. 2003;422:307–312. - PubMed
-
- Burton DR, et al. Efficient neutralization of primary isolates of HIV-1 by a recombinant human monoclonal antibody. Science. 1994;266:1024–1027. - PubMed
-
- Cardoso RM, et al. Broadly neutralizing anti-HIV antibody 4E10 recognizes a helical conformation of a highly conserved fusion-associated motif in gp41. Immunity. 2005;22:163–173. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
