

Cohesion group approach for evolutionary analysis of TyrA, a protein family with wide-ranging substrate specificities
- PMID: 18322033
- PMCID: PMC2268282
- DOI: 10.1128/MMBR.00026-07
Cohesion group approach for evolutionary analysis of TyrA, a protein family with wide-ranging substrate specificities
Abstract
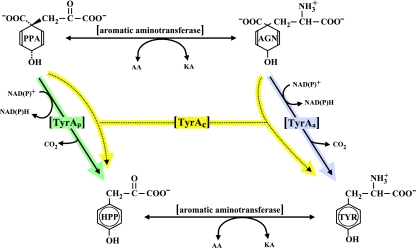
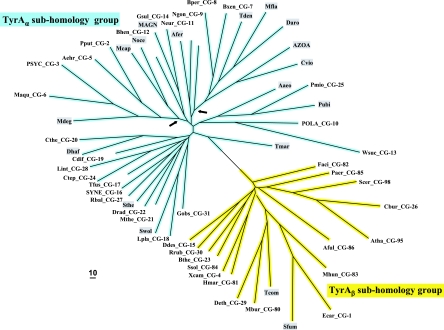
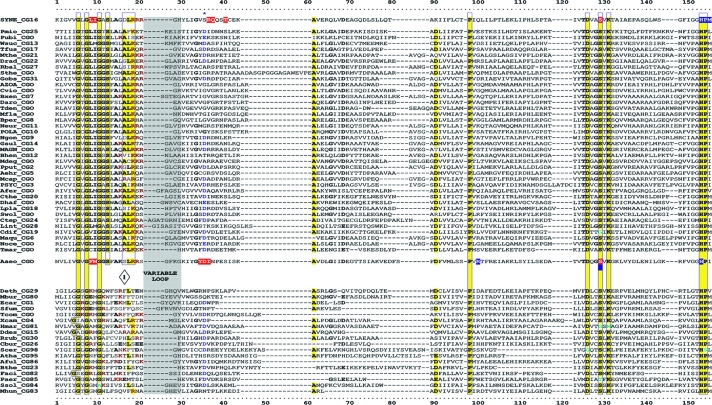
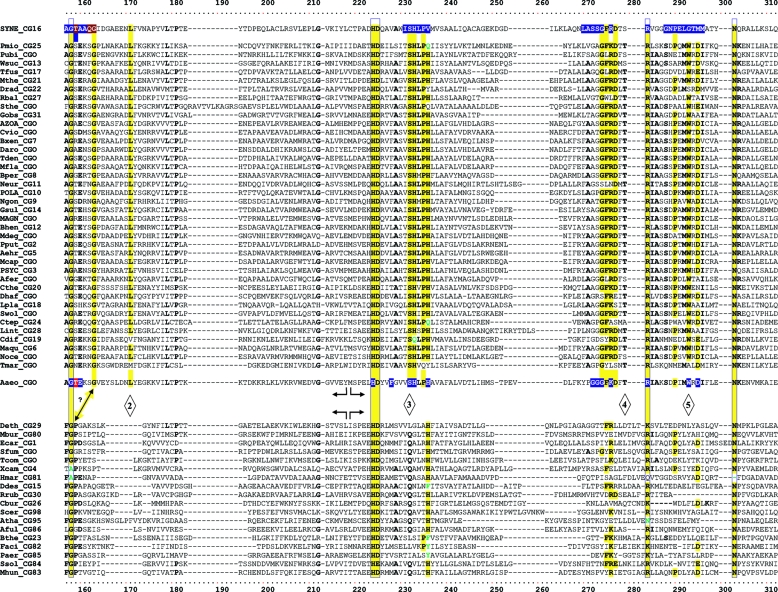
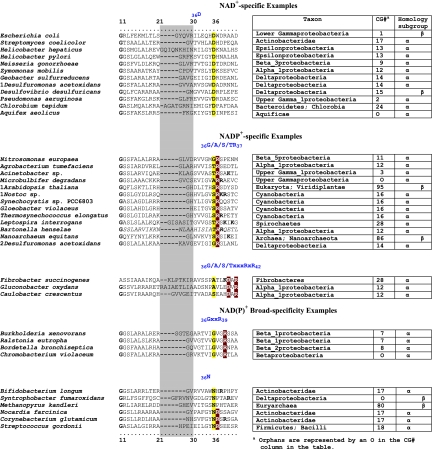
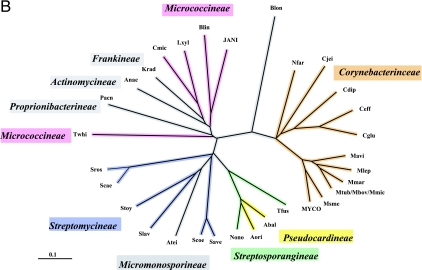
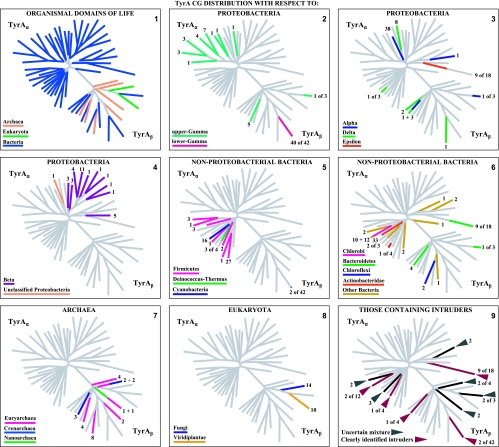
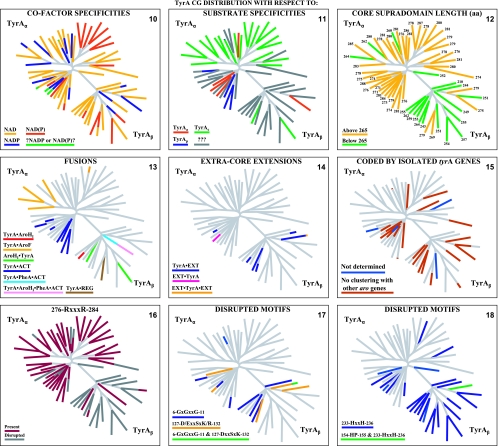
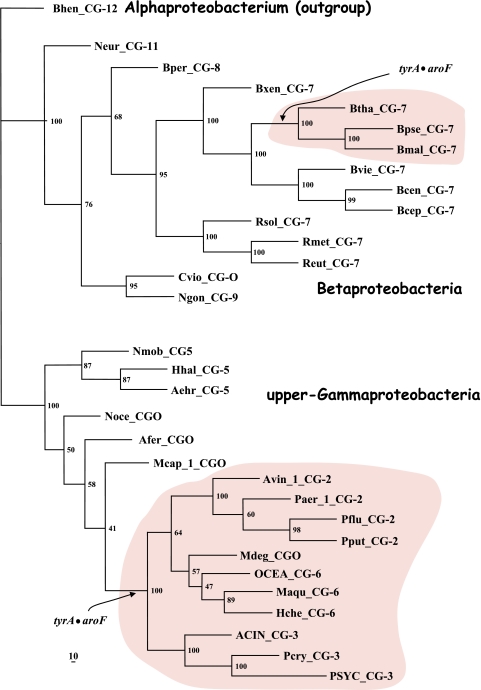
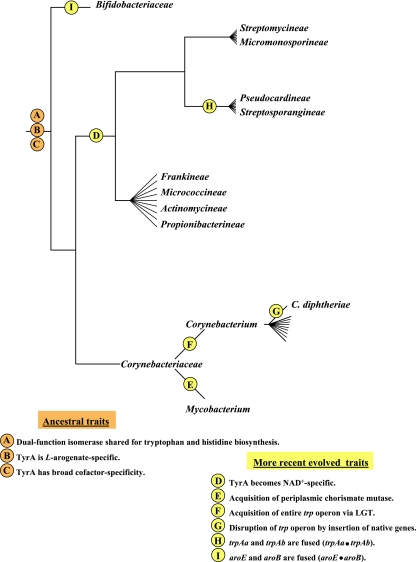
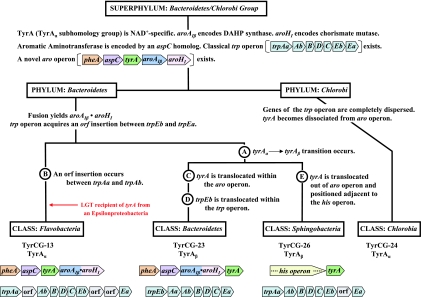
Many enzymes and other proteins are difficult subjects for bioinformatic analysis because they exhibit variant catalytic, structural, regulatory, and fusion mode features within a protein family whose sequences are not highly conserved. However, such features reflect dynamic and interesting scenarios of evolutionary importance. The value of experimental data obtained from individual organisms is instantly magnified to the extent that given features of the experimental organism can be projected upon related organisms. But how can one decide how far along the similarity scale it is reasonable to go before such inferences become doubtful? How can a credible picture of evolutionary events be deduced within the vertical trace of inheritance in combination with intervening events of lateral gene transfer (LGT)? We present a comprehensive analysis of a dehydrogenase protein family (TyrA) as a prototype example of how these goals can be accomplished through the use of cohesion group analysis. With this approach, the full collection of homologs is sorted into groups by a method that eliminates bias caused by an uneven representation of sequences from organisms whose phylogenetic spacing is not optimal. Each sufficiently populated cohesion group is phylogenetically coherent and defined by an overall congruence with a distinct section of the 16S rRNA gene tree. Exceptions that occasionally are found implicate a clearly defined LGT scenario whereby the recipient lineage is apparent and the donor lineage of the gene transferred is localized to those organisms that define the cohesion group. Systematic procedures to manage and organize otherwise overwhelming amounts of data are demonstrated.
Figures

References
-
- Afriat, L., C. Roodveldt, G. Manco, and D. S. Tawfik. 2006. The latent promiscuity of newly identified microbial lactonases is linked to a recently diverged phosphotriesterase. Biochemistry 4513677-13686. - PubMed
-
- Aharoni, A., L. Gaidukov, O. Khersonsky, S. M. Gould, C. Roodveldt, and D. S. Tawfik. 2005. The ‘evolvability’ of promiscuous protein functions. Nat. Genet. 3773-76. - PubMed
-
- Ahmad, S., and R. A. Jensen. 1988. The phylogenetic origin of the bifunctional tyrosine-pathway protein in the enteric lineage of bacteria. Mol. Biol. Evol. 5282-297. - PubMed
-
- Ahmad, S., and R. A. Jensen. 1987. The prephenate dehydrogenase component of the bifunctional T-protein in enteric bacteria can utilize L-arogenate. FEBS Lett. 216133-139. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
