

Mitotic spindle destabilization and genomic instability in Shwachman-Diamond syndrome
- PMID: 18324336
- PMCID: PMC2263145
- DOI: 10.1172/JCI33764
Mitotic spindle destabilization and genomic instability in Shwachman-Diamond syndrome
Abstract
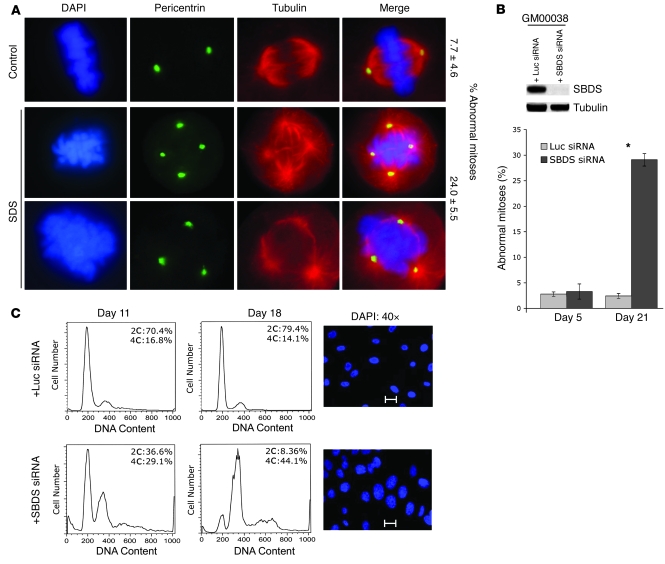
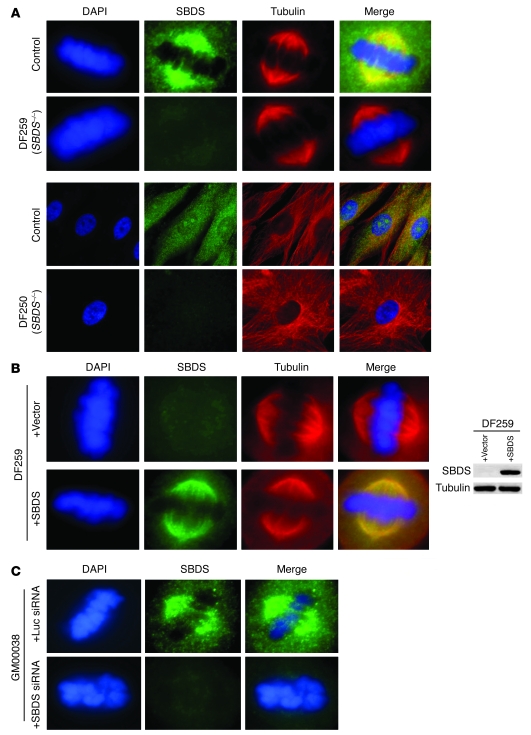
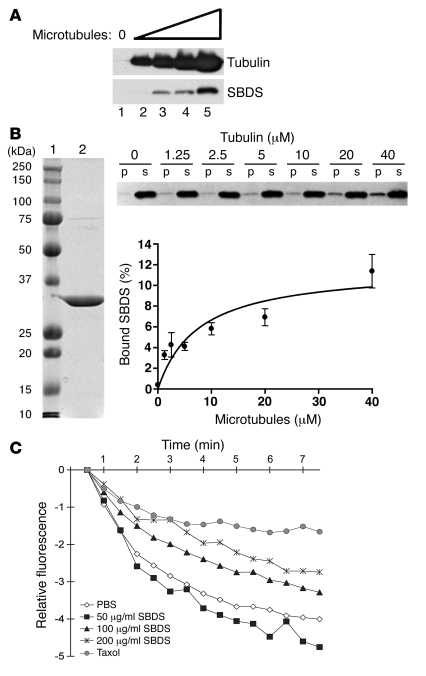
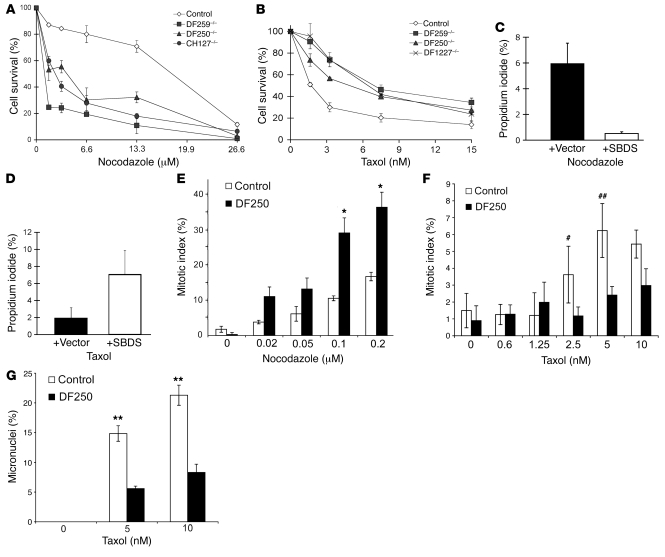
Deficiencies in the SBDS gene result in Shwachman-Diamond syndrome (SDS), an inherited bone marrow failure syndrome associated with leukemia predisposition. SBDS encodes a highly conserved protein previously implicated in ribosome biogenesis. Using human primary bone marrow stromal cells (BMSCs), lymphoblasts, and skin fibroblasts, we show that SBDS stabilized the mitotic spindle to prevent genomic instability. SBDS colocalized with the mitotic spindle in control primary BMSCs, lymphoblasts, and skin fibroblasts and bound to purified microtubules. Recombinant SBDS protein stabilized microtubules in vitro. We observed that primary BMSCs and lymphoblasts from SDS patients exhibited an increased incidence of abnormal mitoses. Similarly, depletion of SBDS by siRNA in human skin fibroblasts resulted in increased mitotic abnormalities and aneuploidy that accumulated over time. Treatment of primary BMSCs and lymphoblasts from SDS patients with nocodazole, a microtubule destabilizing agent, led to increased mitotic arrest and apoptosis, consistent with spindle destabilization. Conversely, SDS patient cells were resistant to taxol, a microtubule stabilizing agent. These findings suggest that spindle instability in SDS contributes to bone marrow failure and leukemogenesis.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
