

Staphylococcus aureus HrtA is an ATPase required for protection against heme toxicity and prevention of a transcriptional heme stress response
- PMID: 18326576
- PMCID: PMC2395006
- DOI: 10.1128/JB.01921-07
Staphylococcus aureus HrtA is an ATPase required for protection against heme toxicity and prevention of a transcriptional heme stress response
Abstract
During systemic infection, Staphylococcus aureus acquires nutrient iron from heme, the cofactor of vertebrate myoglobin and hemoglobin. Upon exposure to heme, S. aureus up-regulates the expression of the heme-regulated transporter, HrtAB. Strains lacking hrtAB exhibit increased sensitivity to heme toxicity, and upon heme exposure they elaborate a secreted protein response that interferes with the recruitment of neutrophils to the site of infection. Taken together, these results have led to the suggestion that hrtAB encodes an efflux system responsible for relieving the toxic effects of accumulated heme. Here we extend these observations by demonstrating that HrtA is the ATPase component of the HrtAB transport system. We show that HrtA is an Mn(2+)/Mg(2+)-dependent ATPase that functions at an optimal pH of 7.5 and exhibits in vitro temperature dependence uncommon to ABC transporter ATPases. Furthermore, we identify conserved residues within HrtA that are required for in vitro ATPase activity and are essential for the functionality of HrtA in vivo. Finally, we show that heme induces an alteration in the gene expression pattern of S. aureus Delta hrtA, implying the presence of a novel transcriptional regulatory mechanism responsible for the previously described immunomodulatory characteristics of hrtA mutants exposed to heme.
Figures
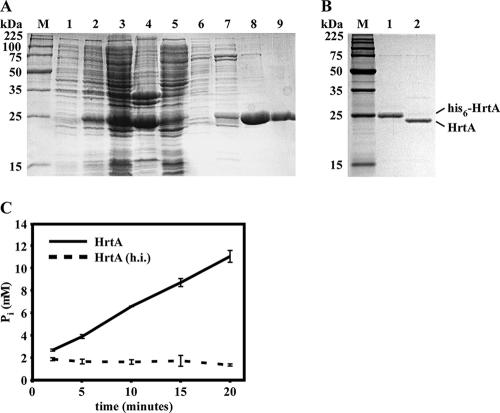
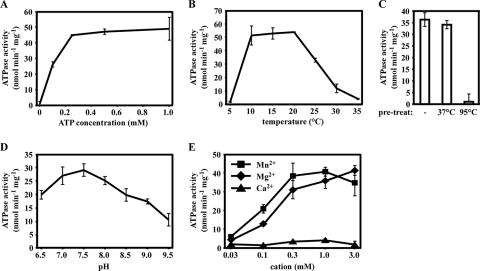
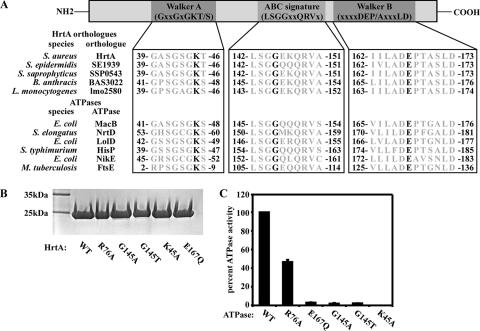
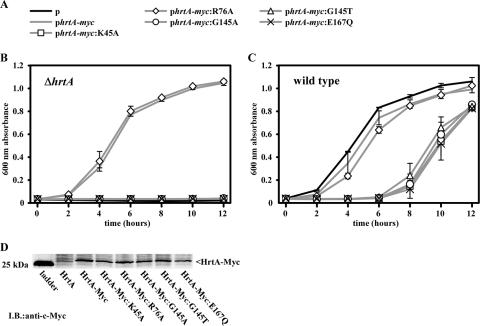
References
-
- Anderson, K. L., C. Roberts, T. Disz, V. Vonstein, K. Hwang, R. Overbeek, P. D. Olson, S. J. Projan, and P. M. Dunman. 2006. Characterization of the Staphylococcus aureus heat shock, cold shock, stringent, and SOS responses and their effects on log-phase mRNA turnover. J. Bacteriol. 1886739-6756. - PMC - PubMed
-
- Borst, P., and R. O. Elferink. 2002. Mammalian ABC transporters in health and disease. Annu. Rev. Biochem. 71537-592. - PubMed
-
- Bullen, J. J., and E. Griffiths. 1999. Iron and infection: molecular, physiological and clinical aspects, vol. John Wiley and Sons, New York, NY.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
