

Neural correlates of perceptual learning in a sensory-motor, but not a sensory, cortical area
- PMID: 18327253
- PMCID: PMC2424192
- DOI: 10.1038/nn2070
Neural correlates of perceptual learning in a sensory-motor, but not a sensory, cortical area
Abstract
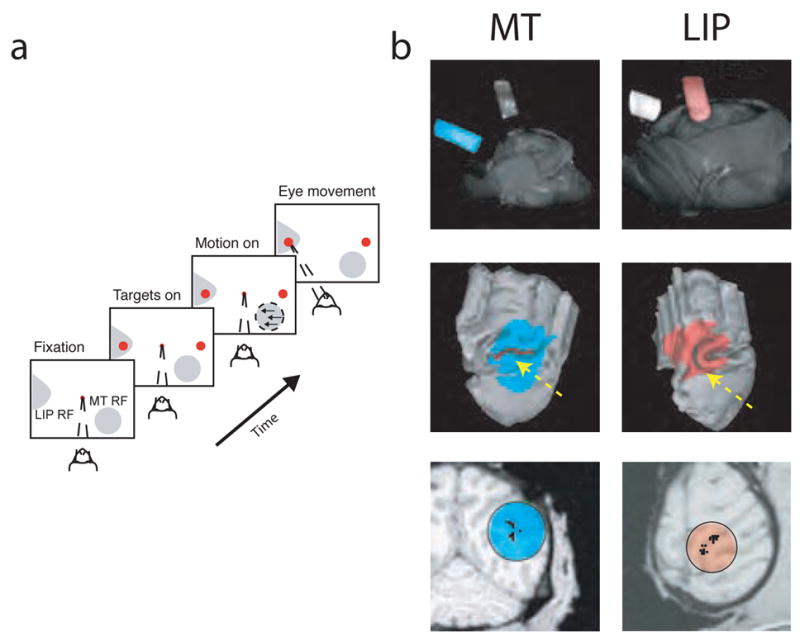
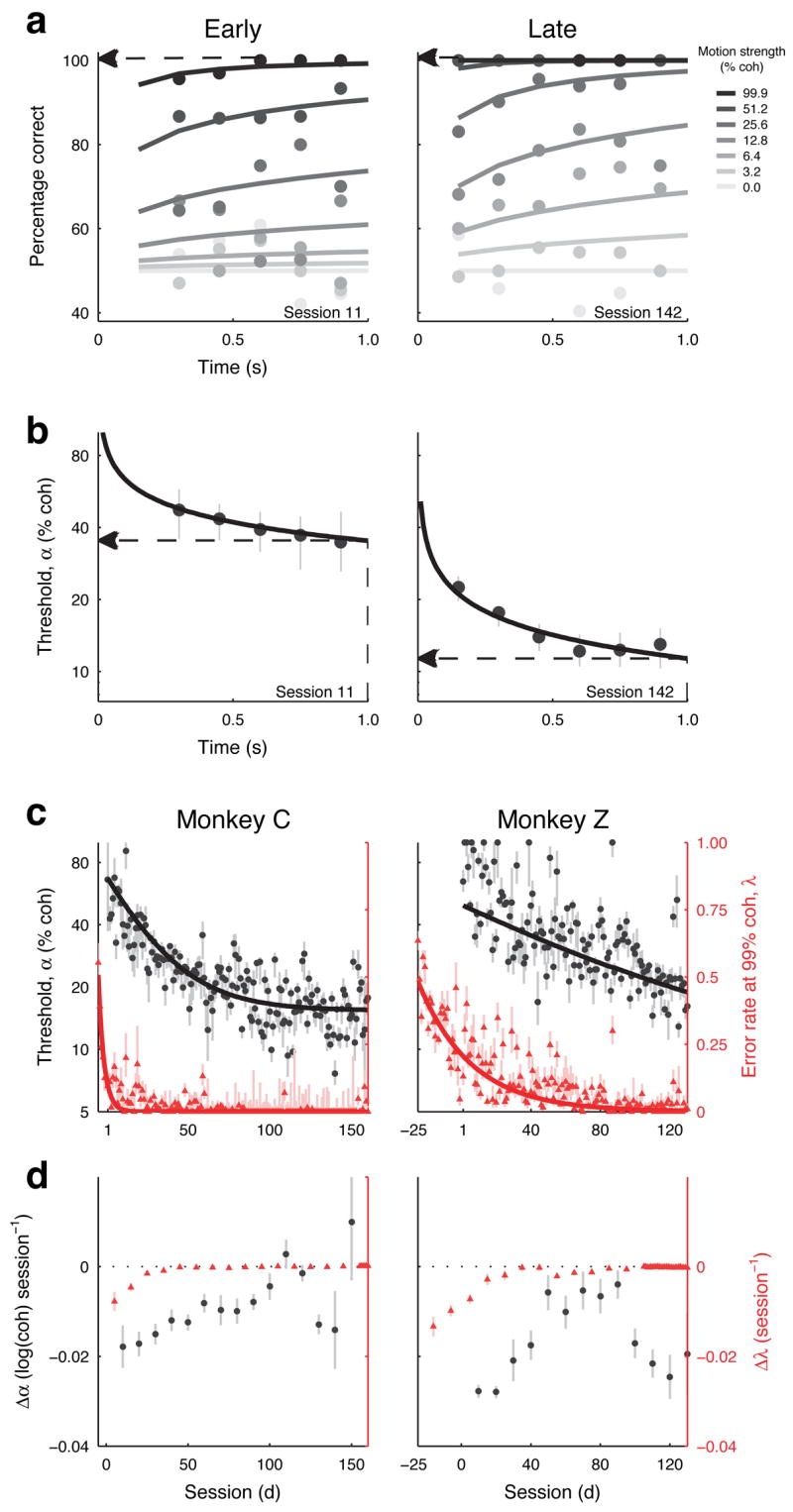
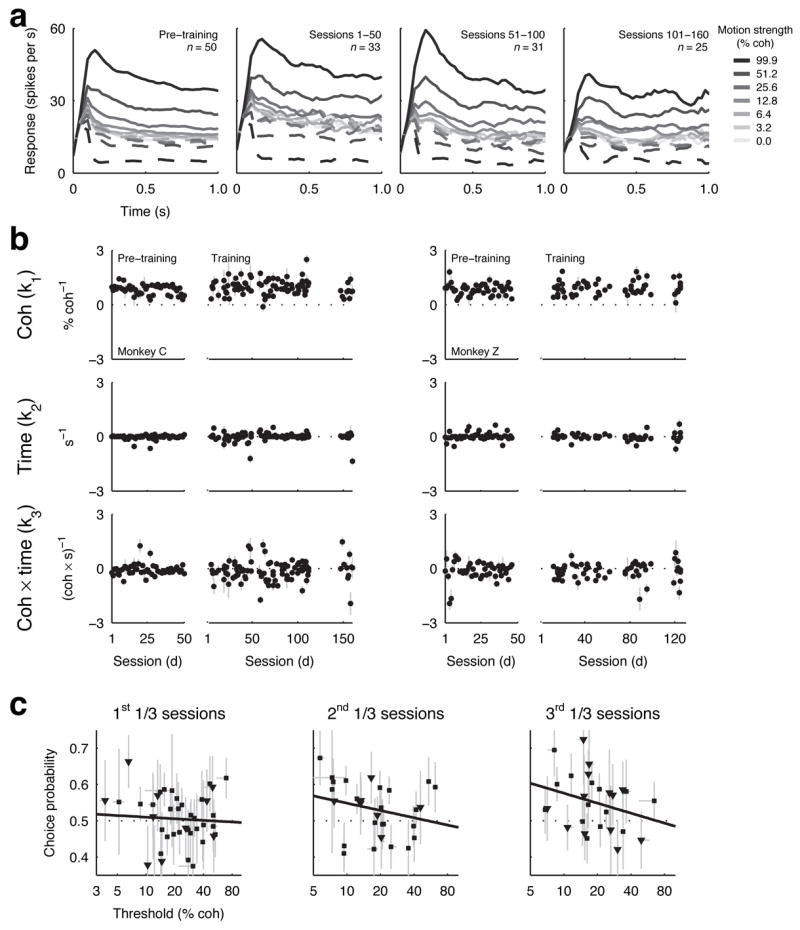
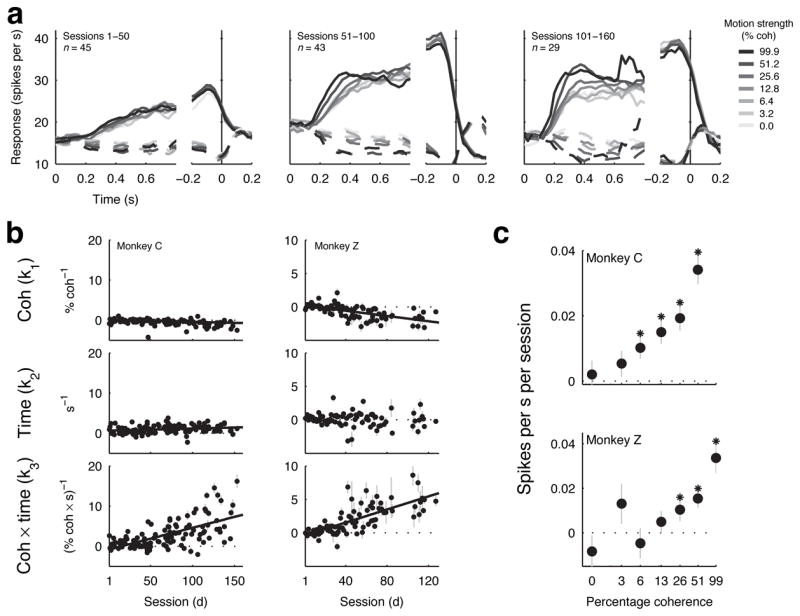
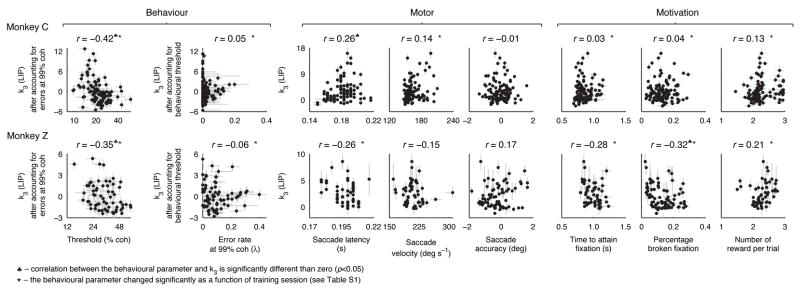
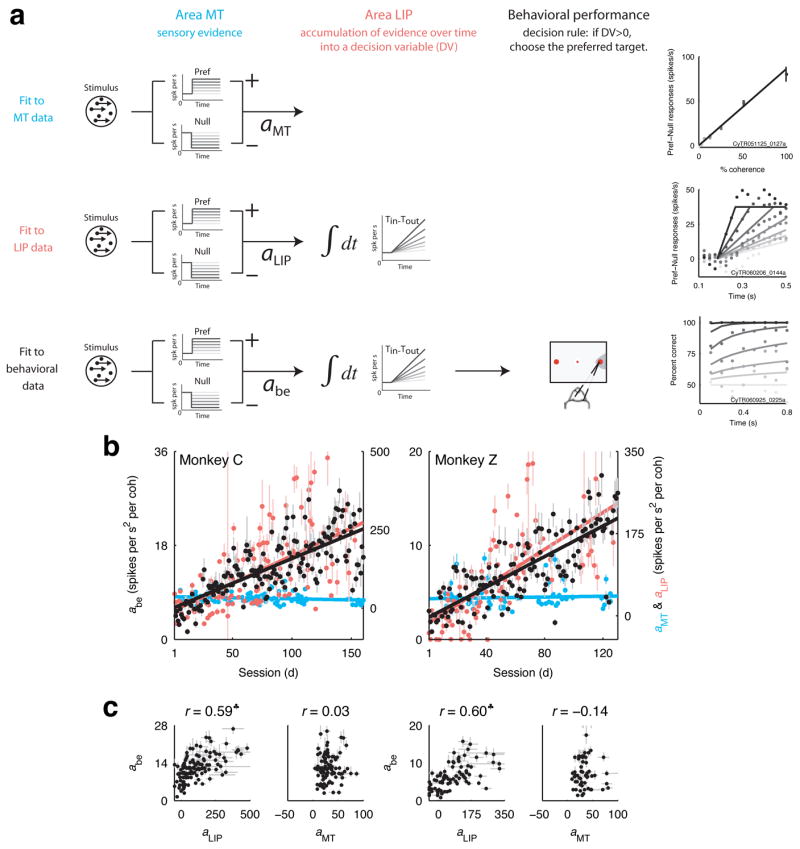
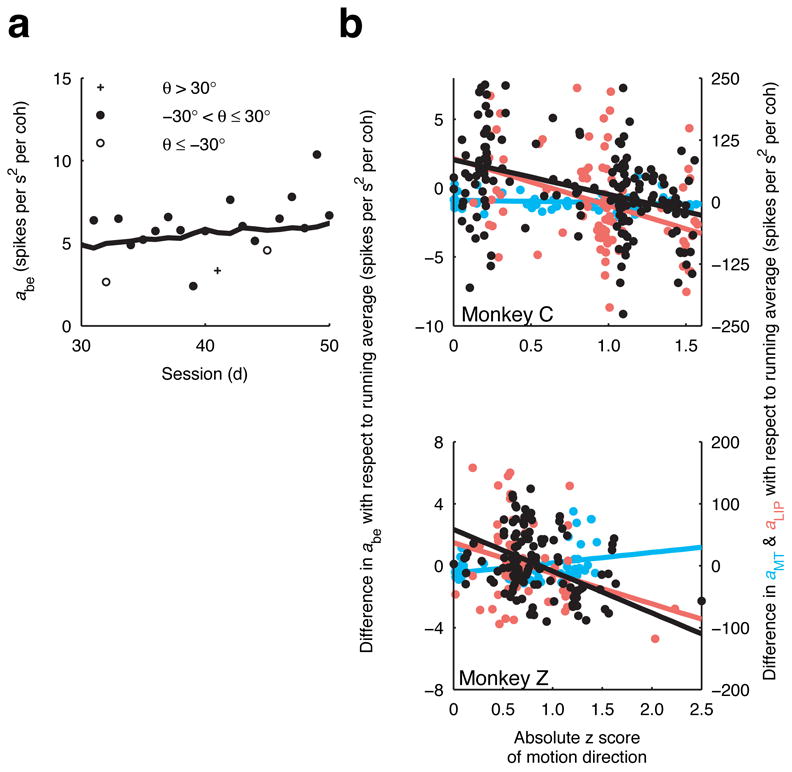
This study aimed to identify neural mechanisms that underlie perceptual learning in a visual-discrimination task. We trained two monkeys (Macaca mulatta) to determine the direction of visual motion while we recorded from their middle temporal area (MT), which in trained monkeys represents motion information that is used to solve the task, and lateral intraparietal area (LIP), which represents the transformation of motion information into a saccadic choice. During training, improved behavioral sensitivity to weak motion signals was accompanied by changes in motion-driven responses of neurons in LIP, but not in MT. The time course and magnitude of the changes in LIP correlated with the changes in behavioral sensitivity throughout training. Thus, for this task, perceptual learning does not appear to involve improvements in how sensory information is represented in the brain, but rather how the sensory representation is interpreted to form the decision that guides behavior.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
