

Pepper pectin methylesterase inhibitor protein CaPMEI1 is required for antifungal activity, basal disease resistance and abiotic stress tolerance
- PMID: 18327607
- PMCID: PMC2413075
- DOI: 10.1007/s00425-008-0719-z
Pepper pectin methylesterase inhibitor protein CaPMEI1 is required for antifungal activity, basal disease resistance and abiotic stress tolerance
Abstract
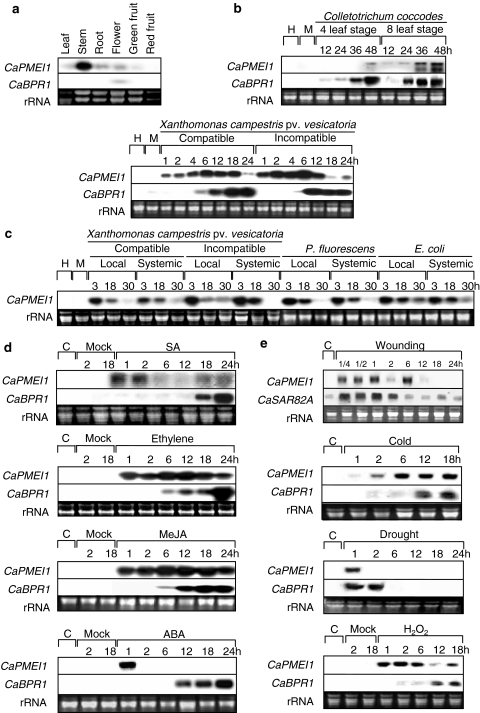
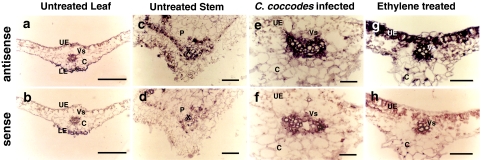
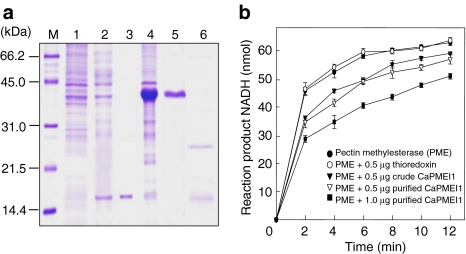
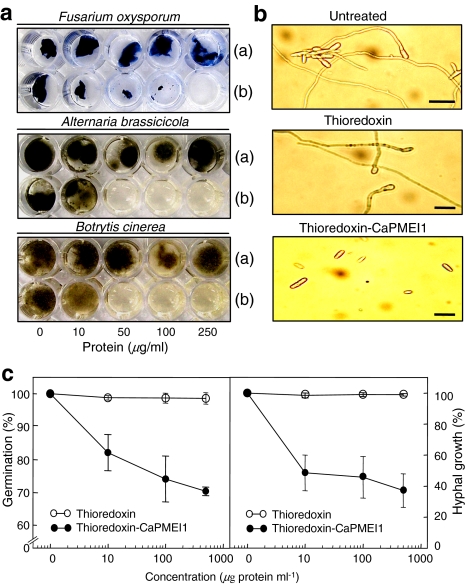
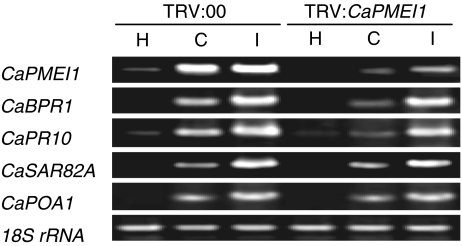
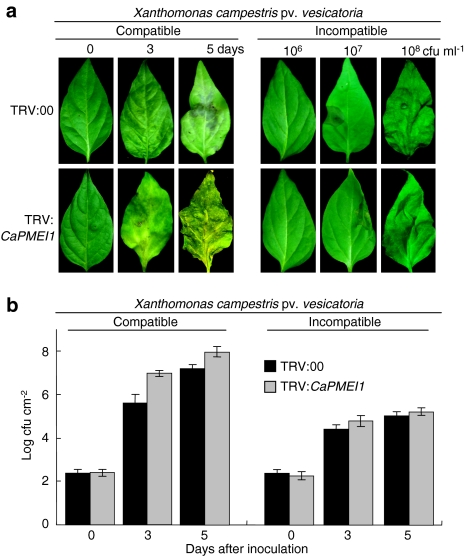
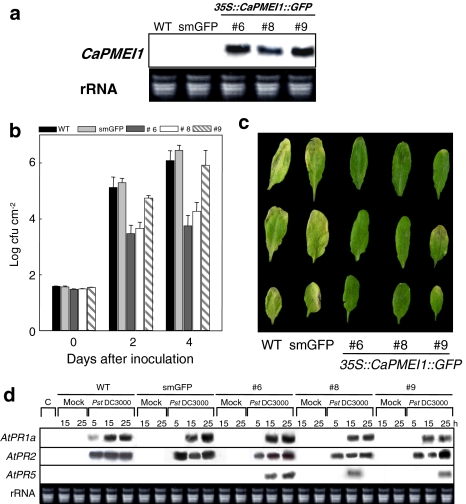
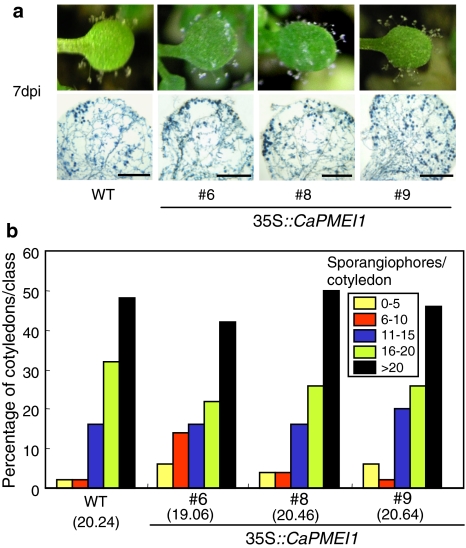
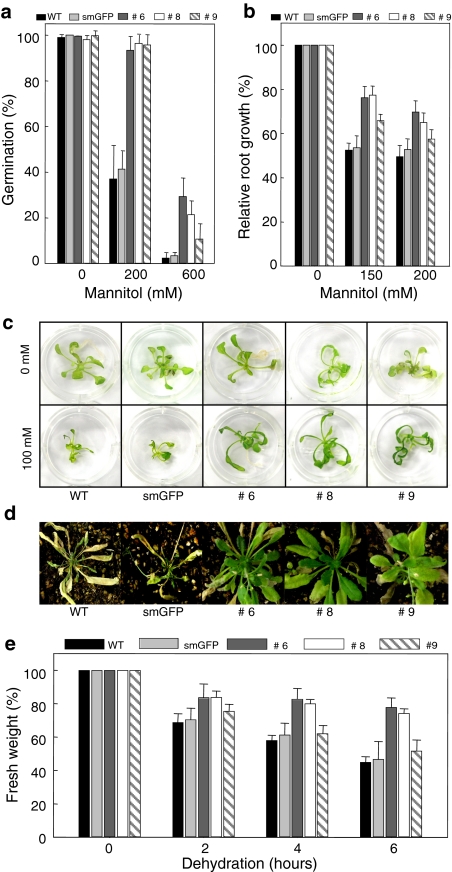
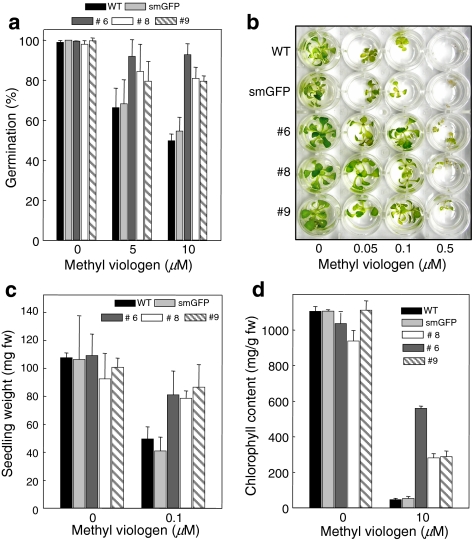
Pectin is one of the main components of the plant cell wall that functions as the primary barrier against pathogens. Among the extracellular pectinolytic enzymes, pectin methylesterase (PME) demethylesterifies pectin, which is secreted into the cell wall in a highly methylesterified form. Here, we isolated and functionally characterized the pepper (Capsicum annuum L.) gene CaPMEI1, which encodes a pectin methylesterase inhibitor protein (PMEI), in pepper leaves infected by Xanthomonas campestris pv. vesicatoria (Xcv). CaPMEI1 transcripts are localized in the xylem of vascular bundles in leaf tissues, and pathogens and abiotic stresses can induce differential expression of this gene. Purified recombinant CaPMEI1 protein not only inhibits PME, but also exhibits antifungal activity against some plant pathogenic fungi. Virus-induced gene silencing of CaPMEI1 in pepper confers enhanced susceptibility to Xcv, accompanied by suppressed expression of some defense-related genes. Transgenic Arabidopsis CaPMEI1-overexpression lines exhibit enhanced resistance to Pseudomonas syringae pv. tomato, mannitol and methyl viologen, but not to the biotrophic pathogen Hyaloperonospora parasitica. Together, these results suggest that CaPMEI1, an antifungal protein, may be involved in basal disease resistance, as well as in drought and oxidative stress tolerance in plants.
Figures

References
-
- Asada K (1996) Radical production and scavenging in the chloroplasts. In: Baker NR (ed) Photosynthesis and the environment. Kluwer, Netherlands, pp. 123–150
-
- Asoufi H, Hameed KM, Mahasneh A (2007) The cellulase and pectinase activities associated with the virulence of indigenous Sclerotinia sclerotiorum isolates in Jordan Valley. Plant Pathol J 23:233–238
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
