

Locally produced complement fragments C5a and C3a provide both costimulatory and survival signals to naive CD4+ T cells
- PMID: 18328742
- PMCID: PMC2646383
- DOI: 10.1016/j.immuni.2008.02.001
Locally produced complement fragments C5a and C3a provide both costimulatory and survival signals to naive CD4+ T cells
Abstract
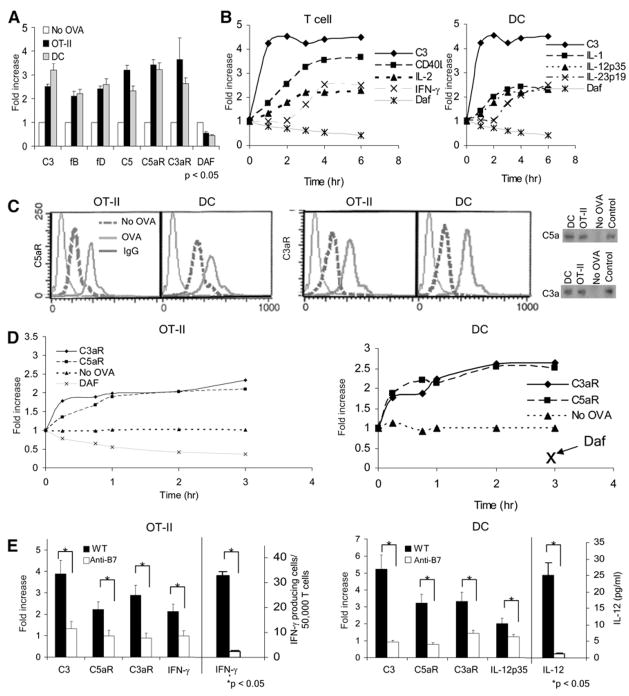
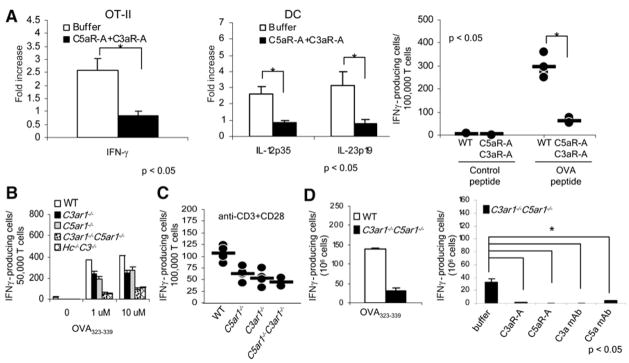
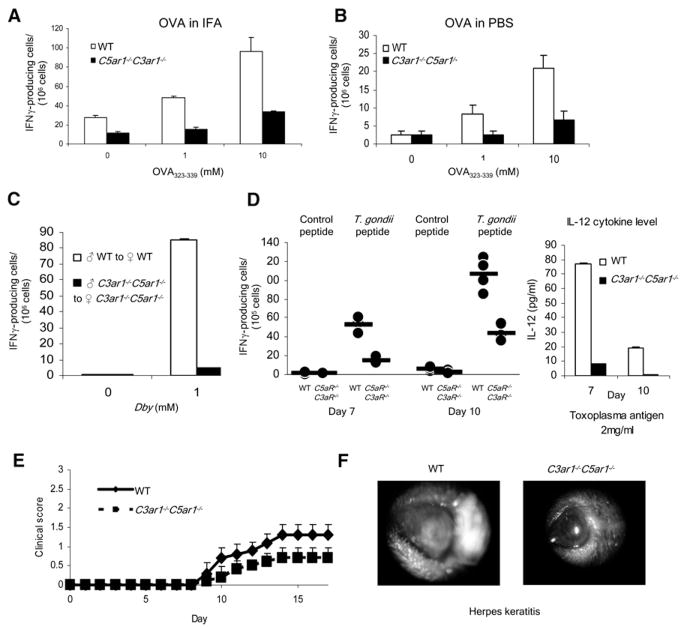
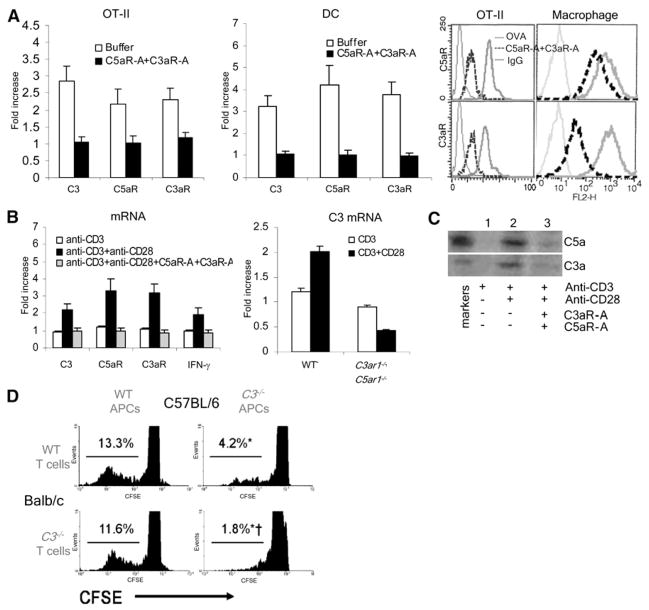
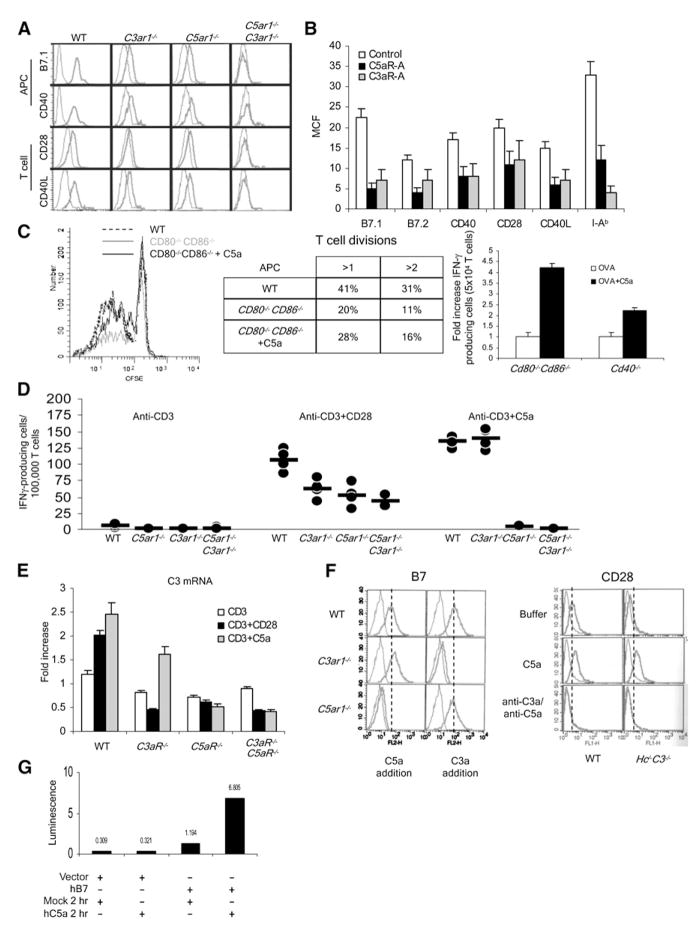
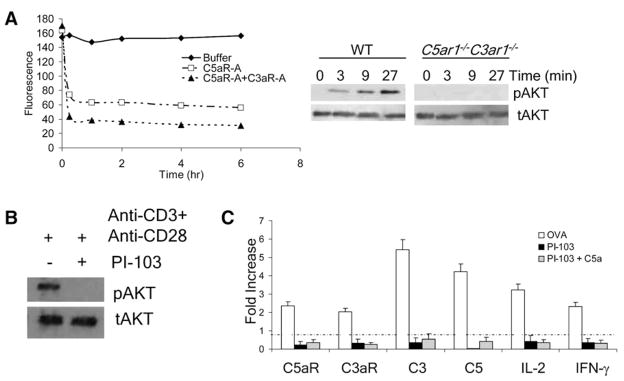
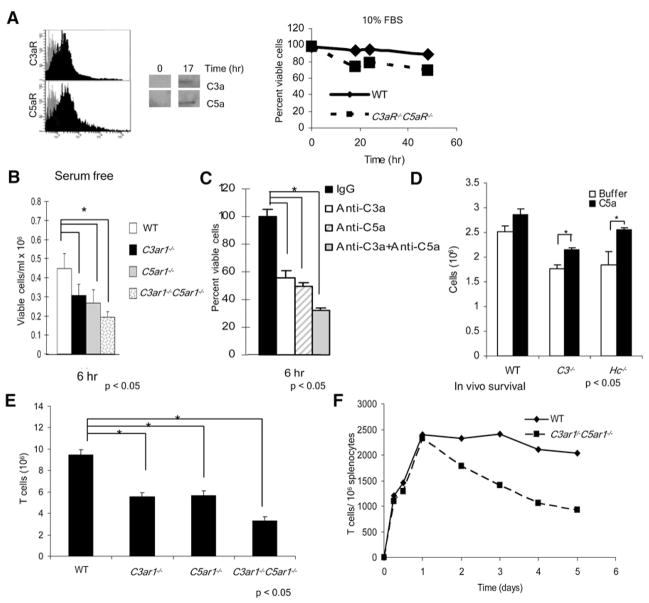
Costimulatory signals are critical to T cell activation, but how their effects are mediated remains incompletely characterized. Here, we demonstrate that locally produced C5a and C3a anaphylatoxins interacting with their G protein-coupled receptors (GPCRs), C5aR and C3aR, on APCs and T cells both upstream and downstream of CD28 and CD40L signaling are integrally involved in T cell proliferation and differentiation. Disabling these interactions reduced MHC class II and costimulatory-molecule expression and dramatically diminished T cell responses. Importantly, impaired T cell activation by Cd80-/-Cd86-/- and Cd40-/- APCs was reconstituted by added C5a or C3a. C5aR and C3aR mediated their effects via PI-3 kinase-gamma-dependent AKT phosphorylation, providing a link between GPCR signaling, CD28 costimulation, and T cell survival. These local paracrine and autocrine interactions thus operate constitutively in naive T cells to maintain viability, and their amplification by cognate APC partners thus is critical to T cell costimulation.
Figures
References
-
- Cain SA, Monk PN. The orphan receptor C5L2 has high affinity binding sites for complement fragments C5a and C5a des-Arg(74) J Biol Chem. 2002;277:7165–7169. - PubMed
-
- Deane JA, Fruman DA. Phosphoinositide 3-kinase: Diverse roles in immune cell activation. Annu Rev Immunol. 2004;22:563–598. - PubMed
-
- Diehl L, Den Boer AT, van der Voort EI, Melief CJ, Offringa R, Toes RE. The role of CD40 in peripheral T cell tolerance and immunity. J Mol Med. 2000;78:363–371. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
