

Comparative Study
doi: 10.1016/0092-8674(91)90043-x.
An evolutionarily conserved cyclin homolog from Drosophila rescues yeast deficient in G1 cyclins
Affiliations
- PMID: 1833067
- PMCID: PMC2753436
- DOI: 10.1016/0092-8674(91)90043-x
Item in Clipboard
Comparative Study
An evolutionarily conserved cyclin homolog from Drosophila rescues yeast deficient in G1 cyclins
Cell.
.
Abstract
We have isolated two Drosophila cDNA clones that rescue Saccharomyces cerevisiae deficient in CLN functions. One of these clones is the Drosophila homolog of the cdc2 gene. The second encodes a distant and new member of the cyclin family of proteins, cyclin C. It is highly homologous (72% identity) to a human clone isolated in a similar screen. Yeast cells rescued by a plasmid constitutively expressing this Drosophila cyclin C are unusually small, consistent with an unregulated high level of G1 cyclin function. Sequence comparisons identified regions conserved among the more distantly related cyclins. Based on these conserved elements, we identified homology between cyclins and the ras oncogene.
Figures
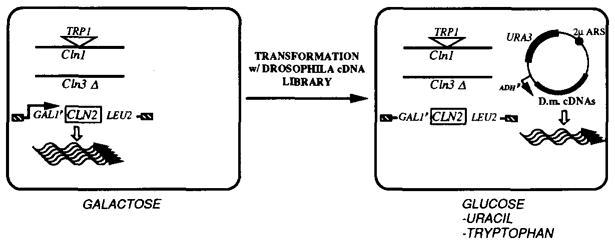
A Drosophila cDNA library in a URA3-marked plasmid was used to transform a strain harboring a deletion of CLN3, a TRP1-disruption of CLN1, and a chromosomal replacement of CLN2 by a GAL1p-driven CLN2 gene. This strain has a triple cln− phenotype when shifted from galactose to glucose media. Cells that contain an introduced plasmid harboring a Drosophila gene providing CLN function can be selected on glucose minimal plates minus uracil. The selection for TRP+ cells was added in order to eliminate the cln+ background, owing to gene conversion between CLN1 and CLN2 sequences.

The nucleotide sequence (1086 bp) has an open reading frame encoding a 267 amino acid protein. The putative initiating ATG is underlined along with flanking bases matching the consensus sequences for translational start sites in Drosophila (Cavener, 1987).

The bars in the upper panel are scaled representations of five divergent cyclins. The location of the three regions identified as having high conservation are indicated as shaded segments. The scale is indicated on the right. For each of the conserved regions, a consensus of G2 cyclin sequences is shown. This was derived as follows: 15 G2 cyclin sequences were compared (clam cyclins A [Swenson et al., 1986] and B [Westendorf et al., 1989], human cyclins A [Wang et al., 1990] and B [Pines and Hunter, 1989], Drosophila cyclins A [Lehner and O’Farrell, 1989] and B [Lehner and O’Farrell, 1990], frog cyclins A [Minshull et al., 1990], B1 [Minshull et al., 1989], and B2 [Minshull et al., 1989], sea urchin cyclin [Pines and Hunt, 1987], S. pombe cdc13 [Booher and Beach, 1988], and S. cerevisiae Clb1, Clb2, Clb3, and Clb4 [Ghiara et al., 1991]). At each position, the most commonly occurring amino acid was selected for the consensus. In the few cases of a numerical tie, we selected the residue that was the most widely distributed among the more evolutionarily diverged cyclins. Below the consensus, all of the different residues that occur at a position are listed as alternatives. The cyclin C sequence and the other diverged cyclin sequences were aligned with the G2 consensus sequence. The number to the left of each aligned sequence is the position of the first residue of the segment. As a reference and an example of a G2 cyclin, the clam cyclin A sequence is included in this alignment. Residues matching the G2 consensus are boxed and shaded. Residues considered homologous to the residues of the consensus are boxed, according to the following groupings: A, L, V, I, M; D, E; K, R; N, Q; F, Y; S, T. The scores on the right of each alignment were calculated as follows. The 15 G2 sequences were used as a reference. The number of times each amino acid occurred at a given position in this reference set was determined. For an aligned sequence, each position was given a score corresponding to the number of times the corresponding amino acid occurred in the reference set. The total scores for an alignment were then normalized by dividing by the maximum score (number of positions scored × number of sequences in the reference set). For the ras homology within region III, homology scores were similarly calculated based only on the 21 amino acid sequence indicated for human H-ras.

Total Drosophila and human sequences are aligned. Shaded boxes correspond to identical residues. The dash indicates a 1 amino acid gap introduced into the human sequence.
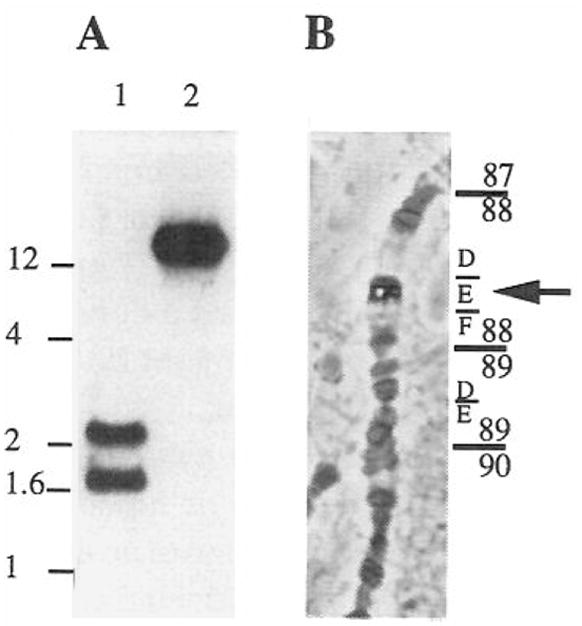
(A) Total genomic DNA from Drosophila was digested with EcoRI (lane 1) and Pstl (lane 2) and probed on Southern blots with a DmcycC probe. The scale on the left is in kilobases. (B) A DmcycC probe was used to localize the gene on polytene chromosomes from salivary glands of third instar larvae. The cyclin C cDNA clone was labeled with digoxigenin and hybridized to salivary gland squashes. The position of the phosphatase signal following immunological detection was identified in several squashes. It consistently mapped to 88E.

Total RNA samples (corresponding to: lane 1, rapid cleavage stage embryos; lane 2, embryos in interphase of cycle 14; lane 3, embryos engaged in divisions 15 and 16; lane 4, older embryos, whose cell divisions are restricted to the nervous system) were probed with the cyclin C cDNA clone on a Northern blot. The lower panel shows a control hybridization of the same filter with an actin 5C probe. Note that the second lane is shifted down owing to loading of an unusually large volume of sample and that cyclin C and actin bands are affected similarly. Given the specific activities of the cyclin C and actin probes and the exposure times of the two hybridizations, we estimate that the intensity of the cyclin C signal is about 200 times less than that of actin. Signals were detected and analyzed by a Molecular Dynamics Phospholmager, which produces a relatively grainy image. The estimated size of the cyclin C mRNA is indicated in kilobases. Embryos were collected and aged at 25°C for the times indicated.
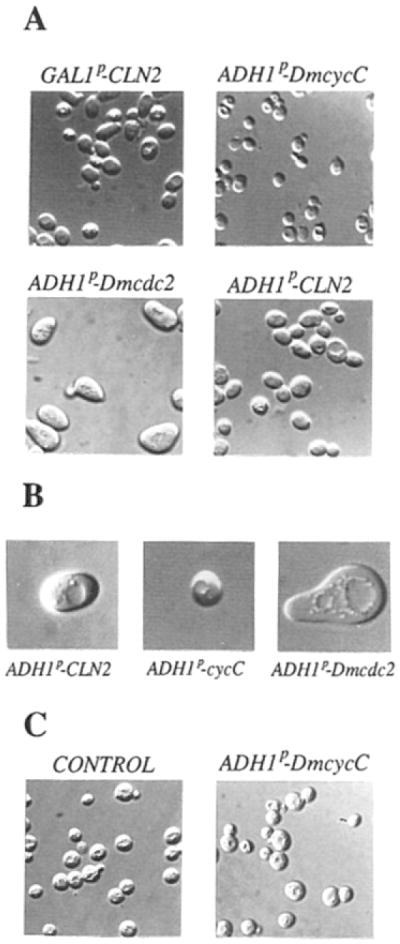
(A) Haploid YPL1 strain grown either in galactose (GAL1p–CLN2) or in glucose after transformation with plasmids containing ADH1 promoter–driven DmcycC, Dmcdc2, or CLN2 (respectively, ADH1p–DmcycC, ADH1p–Dmcdc2, and ADH1p–CLN2). (B) Enlargements of representative YPL1 transformants with ADH1p–CLN2, ADHIp–DmcycC, and ADH1p–Dmcdc2. (C) CLN+(bar1-1) cells either transformed with ADH1p–DmcycC or non-transformed (CONTROL), grown in glucose selective media minus uracil, or supplemented with uracil in the case of the nontransformed control.
References
-
- Burgers PM, Percival KJ. Transformation of yeast spheroplasts without cell fusion. Anal Biochem. 1987;163:391–397. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
