

MURC, a muscle-restricted coiled-coil protein that modulates the Rho/ROCK pathway, induces cardiac dysfunction and conduction disturbance
- PMID: 18332105
- PMCID: PMC2423172
- DOI: 10.1128/MCB.02186-07
MURC, a muscle-restricted coiled-coil protein that modulates the Rho/ROCK pathway, induces cardiac dysfunction and conduction disturbance
Abstract
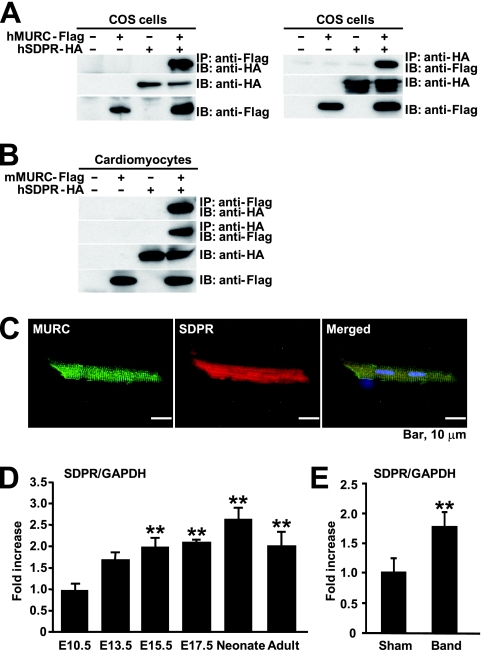
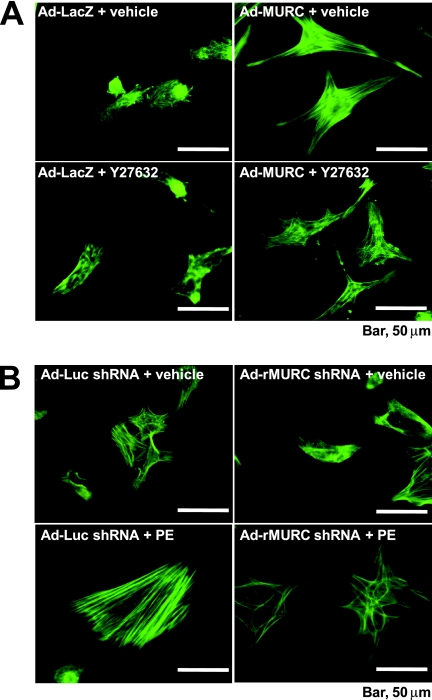
We identified a novel muscle-restricted putative coiled-coil protein, MURC, which is evolutionarily conserved from frog to human. MURC was localized to the cytoplasm with accumulation in the Z-line of the sarcomere in the murine adult heart. MURC mRNA expression in the heart increased during the developmental process from the embryonic stage to adulthood. In response to pressure overload, MURC mRNA expression increased in the hypertrophied heart. Using the yeast two-hybrid system, we identified the serum deprivation response (SDPR) protein, a phosphatidylserine-binding protein, as a MURC-binding protein. MURC induced activation of the RhoA/ROCK pathway, which modulated serum response factor-mediated atrial natriuretic peptide (ANP) expression and myofibrillar organization. SDPR augmented MURC-induced transactivation of the ANP promoter in cardiomyocytes, and RNA interference of SDPR attenuated the action of MURC on the ANP promoter. Transgenic mice expressing cardiac-specific MURC (Tg-MURC) exhibited cardiac contractile dysfunction and atrioventricular (AV) conduction disturbances with atrial chamber enlargement, reduced thickness of the ventricular wall, and interstitial fibrosis. Spontaneous episodes of atrial fibrillation and AV block were observed in Tg-MURC mice. These findings indicate that MURC modulates RhoA signaling and that MURC plays an important role in the development of cardiac dysfunction and conduction disturbance with increased vulnerability to atrial arrhythmias.
Figures









Similar articles
-
Hypoxia activates muscle-restricted coiled-coil protein (MURC) expression via transforming growth factor-β in cardiac myocytes.Clin Sci (Lond). 2014 Mar;126(5):367-75. doi: 10.1042/CS20130260. Clin Sci (Lond). 2014. PMID: 24001173
-
The coiled-coil domain of MURC/cavin-4 is involved in membrane trafficking of caveolin-3 in cardiomyocytes.Am J Physiol Heart Circ Physiol. 2015 Dec 15;309(12):H2127-36. doi: 10.1152/ajpheart.00446.2015. Epub 2015 Oct 23. Am J Physiol Heart Circ Physiol. 2015. PMID: 26497963
-
Effect of atorvastatin on cardiomyocyte hypertrophy through suppressing MURC induced by volume overload and cyclic stretch.J Cell Mol Med. 2019 Feb;23(2):1406-1414. doi: 10.1111/jcmm.14044. Epub 2018 Dec 3. J Cell Mol Med. 2019. PMID: 30511410 Free PMC article.
-
Functions of Rho family of small GTPases and Rho-associated coiled-coil kinases in bone cells during differentiation and mineralization.Biochim Biophys Acta Gen Subj. 2017 May;1861(5 Pt A):1009-1023. doi: 10.1016/j.bbagen.2017.02.005. Epub 2017 Feb 8. Biochim Biophys Acta Gen Subj. 2017. PMID: 28188861 Review.
-
Revisited and revised: is RhoA always a villain in cardiac pathophysiology?J Cardiovasc Transl Res. 2010 Aug;3(4):330-43. doi: 10.1007/s12265-010-9192-8. Epub 2010 May 27. J Cardiovasc Transl Res. 2010. PMID: 20559774 Free PMC article. Review.
Cited by
-
The Cardiac Syndecan-2 Interactome.Front Cell Dev Biol. 2020 Aug 28;8:792. doi: 10.3389/fcell.2020.00792. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32984315 Free PMC article.
-
Systems Network Genomic Analysis Reveals Cardioprotective Effect of MURC/Cavin-4 Deletion Against Ischemia/Reperfusion Injury.J Am Heart Assoc. 2019 Aug 6;8(15):e012047. doi: 10.1161/JAHA.119.012047. Epub 2019 Jul 31. J Am Heart Assoc. 2019. PMID: 31364493 Free PMC article.
-
Clathrin-independent pathways of endocytosis.Cold Spring Harb Perspect Biol. 2014 Jun 2;6(6):a016758. doi: 10.1101/cshperspect.a016758. Cold Spring Harb Perspect Biol. 2014. PMID: 24890511 Free PMC article.
-
The human cardiac and skeletal muscle proteomes defined by transcriptomics and antibody-based profiling.BMC Genomics. 2015 Jun 25;16(1):475. doi: 10.1186/s12864-015-1686-y. BMC Genomics. 2015. PMID: 26109061 Free PMC article.
-
FSHD myotubes with different phenotypes exhibit distinct proteomes.PLoS One. 2012;7(12):e51865. doi: 10.1371/journal.pone.0051865. Epub 2012 Dec 18. PLoS One. 2012. PMID: 23272181 Free PMC article.
References
-
- Adam, O., G. Frost, F. Custodis, M. A. Sussman, H. J. Schafers, M. Bohm, and U. Laufs. 2007. Role of Rac1 GTPase activation in atrial fibrillation. J. Am. Coll. Cardiol. 50359-367. - PubMed
-
- Amano, M., Y. Fukata, and K. Kaibuchi. 2000. Regulation and functions of Rho-associated kinase. Exp. Cell. Res. 26144-51. - PubMed
-
- Aoki, H., S. Izumo, and J. Sadoshima. 1998. Angiotensin II activates RhoA in cardiac myocytes: a critical role of RhoA in angiotensin II-induced premyofibril formation. Circ. Res. 82666-676. - PubMed
-
- Arai, A., J. A. Spencer, and E. N. Olson. 2002. STARS, a striated muscle activator of Rho signaling and serum response factor-dependent transcription. J. Biol. Chem. 27724453-24459. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous