

Colicin N binds to the periphery of its receptor and translocator, outer membrane protein F
- PMID: 18334212
- PMCID: PMC2581486
- DOI: 10.1016/j.str.2007.12.023
Colicin N binds to the periphery of its receptor and translocator, outer membrane protein F
Abstract
Colicins kill Escherichia coli after translocation across the outer membrane. Colicin N displays an unusually simple translocation pathway, using the outer membrane protein F (OmpF) as both receptor and translocator. Studies of this binary complex may therefore reveal a significant component of the translocation pathway. Here we show that, in 2D crystals, colicin is found outside the porin trimer, suggesting that translocation may occur at the protein-lipid interface. The major lipid of the outer leaflet interface is lipopolysaccharide (LPS). It is further shown that colicin N binding displaces OmpF-bound LPS. The N-terminal helix of the pore-forming domain, which is not required for pore formation, rearranges and binds to OmpF. Colicin N also binds artificial OmpF dimers, indicating that trimeric symmetry plays no part in the interaction. The data indicate that colicin is closely associated with the OmpF-lipid interface, providing evidence that this peripheral pathway may play a role in colicin transmembrane transport.
Figures
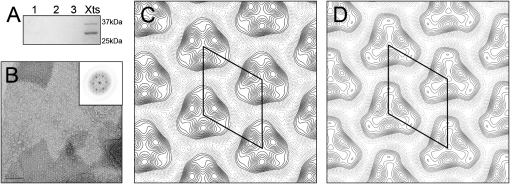
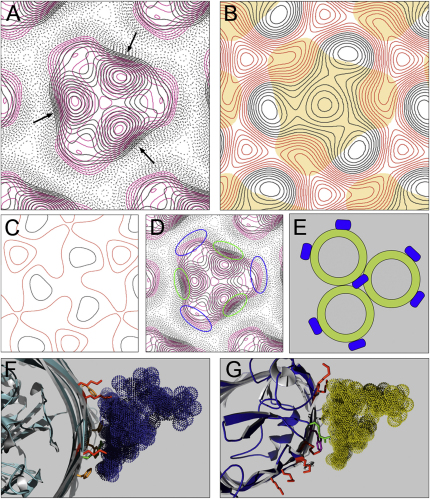
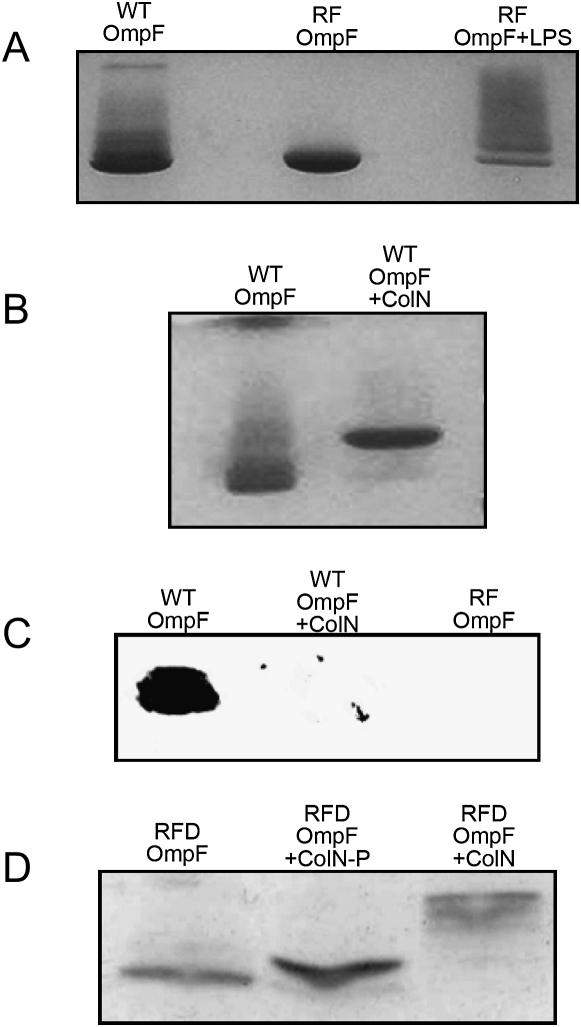
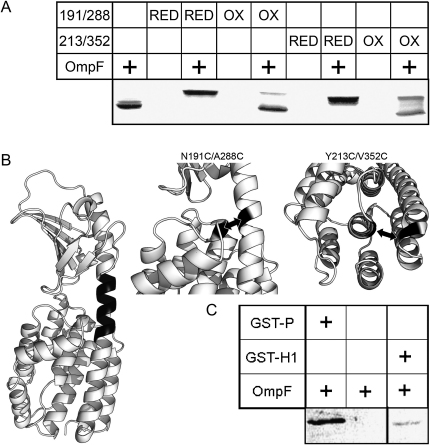
References
-
- Anderluh G., Dalla Serra M., Viero G., Guella G., Macek P., Menestrina G. Pore formation by equinatoxin II, a eukaryotic protein toxin, occurs by induction of nonlamellar lipid structures. J. Biol. Chem. 2003;278:45216–45223. - PubMed
-
- Bainbridge G., Armstrong G.A., Dover L.G., Whelan K.F., Lakey J.H. Displacement of OmpF loop 3 is not required for the membrane translocation of colicins N and A in vivo. FEBS Lett. 1998;432:117–122. - PubMed
-
- Barlic A., Gutierrez-Aguirre I., Caaveiro J.M.M., Cruz A., Ruiz-Arguello M.B., Perez-Gil J., Gonzalez-Manas J.M. Lipid phase coexistence favors membrane insertion of equinatoxin-II, a pore-forming toxin from Actinia equina. J. Biol. Chem. 2004;279:34209–34216. - PubMed
-
- Baty D., Lakey J., Pattus F., Lazdunski C. A 136-amino-acid-residue COOH-terminal fragment of colicin A is endowed with ionophoric activity. Eur. J. Biochem. 1990;189:409–413. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
