

A role for non-rapid-eye-movement sleep homeostasis in perceptual learning
- PMID: 18337406
- PMCID: PMC6670683
- DOI: 10.1523/JNEUROSCI.5548-07.2008
A role for non-rapid-eye-movement sleep homeostasis in perceptual learning
Abstract
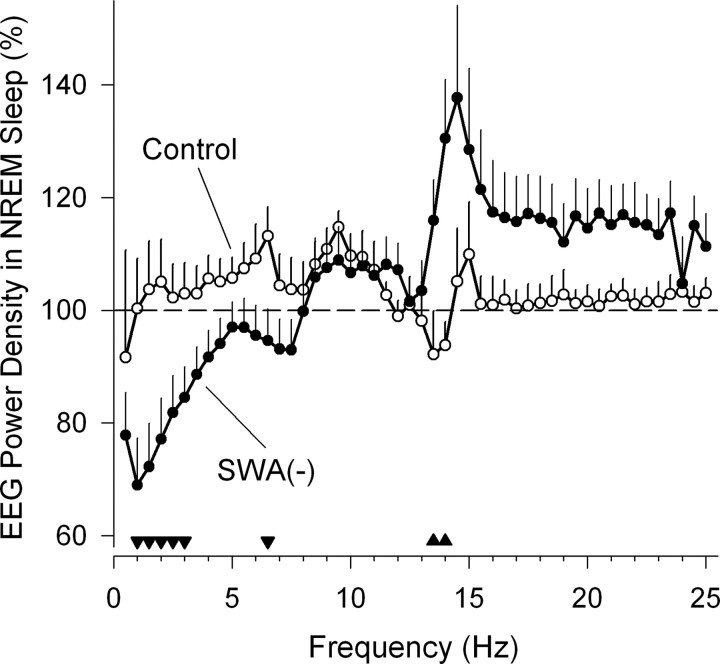
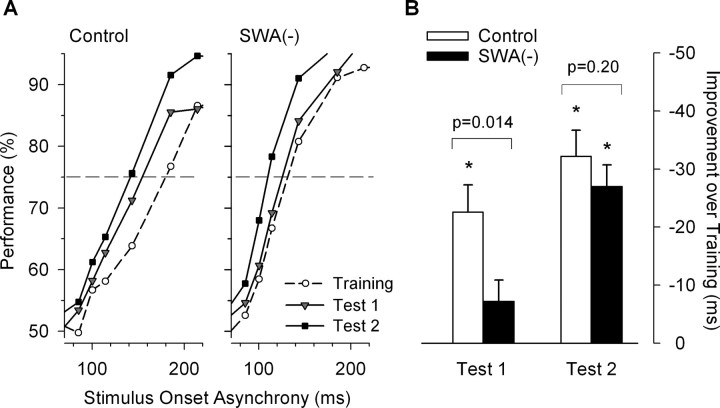
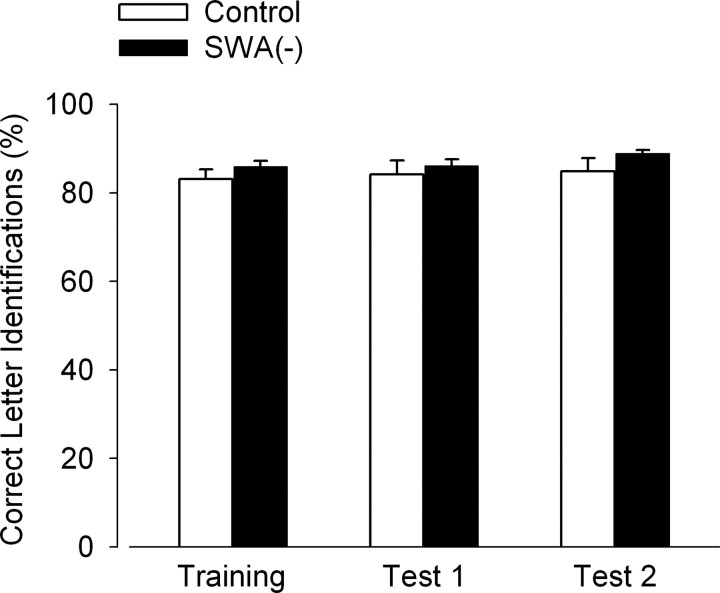
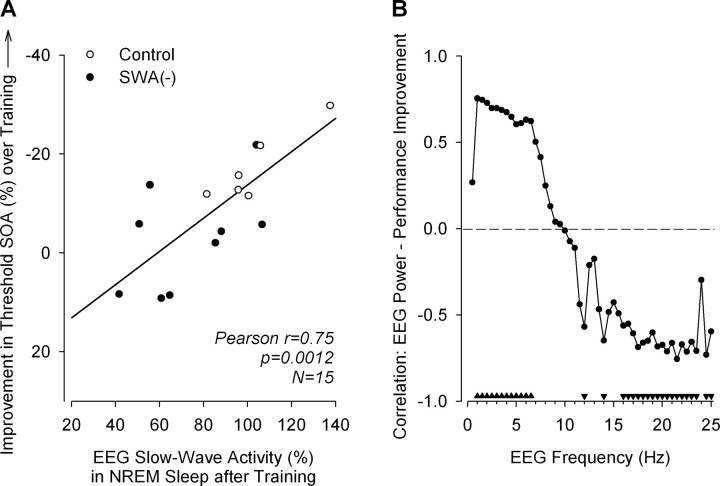
Slow-wave activity (SWA; EEG power density in the 0.75-4.5 Hz range) in non-rapid-eye-movement (NREM) sleep is the primary marker of sleep homeostasis and thought to reflect sleep need. But it is unknown whether the generation of SWA itself serves a fundamental function. Previously, SWA has been implicated in brain plasticity and learning, yet the evidence for a causal role remains correlative. Here, we used acoustic slow-wave suppression to test whether overnight improvement in visual texture discrimination, a form of perceptual learning, directly depends on SWA during sleep. Two groups of subjects were trained on a texture discrimination task (TDT) after baseline sleep, and were tested 24 h later, after a 4 h experimental (EX) sleep episode (with or without SWA suppression), and again after a night of recovery sleep. In the suppression group, SWA during EX sleep was reduced by 30% compared with the control group, whereas total sleep time and REM sleep were not affected. Texture discrimination improved after EX sleep in the control group but not in the suppression group. Moreover, overnight improvement in TDT performance correlated with EEG power density during NREM sleep in the frequency range of SWA (maximum r = 0.75 at 0.75-1.0 Hz) over brain areas involved in TDT learning. We conclude that SWA is an important determinant of sleep-dependent gains in perceptual performance, a finding that directly implicates processes of sleep homeostasis in learning.
Figures
References
-
- Achermann P, Finelli LA, Borbely AA. Unihemispheric enhancement of delta power in human frontal sleep EEG by prolonged wakefulness. Brain Res. 2001;913:220–223. - PubMed
-
- Aeschbach D, Borbély AA. All-night dynamics of the human sleep EEG. J Sleep Res. 1993;2:70–81. - PubMed
-
- Aeschbach D, Cajochen C, Landolt H-P, Borbély AA. Homeostatic sleep regulation in habitual short sleepers and long sleepers. Am J Physiol. 1996;270:R41–R53. - PubMed
-
- Blake H, Gerard RW. Brain potentials during sleep. Am J Physiol. 1937;119:692–703.
-
- Borbély AA. A two process model of sleep regulation. Hum Neurobiol. 1982;1:195–204. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources