

Insulin signaling is involved in the regulation of worker division of labor in honey bee colonies
- PMID: 18337502
- PMCID: PMC2393790
- DOI: 10.1073/pnas.0800630105
Insulin signaling is involved in the regulation of worker division of labor in honey bee colonies
Abstract
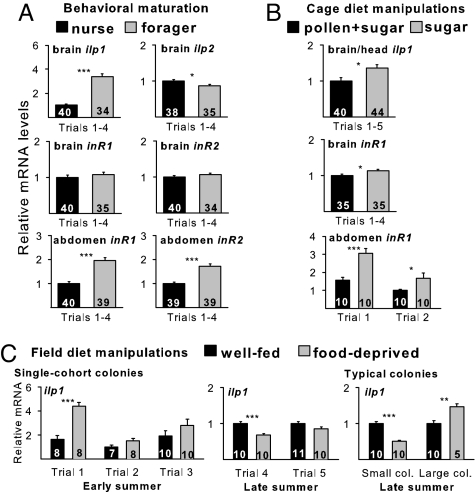
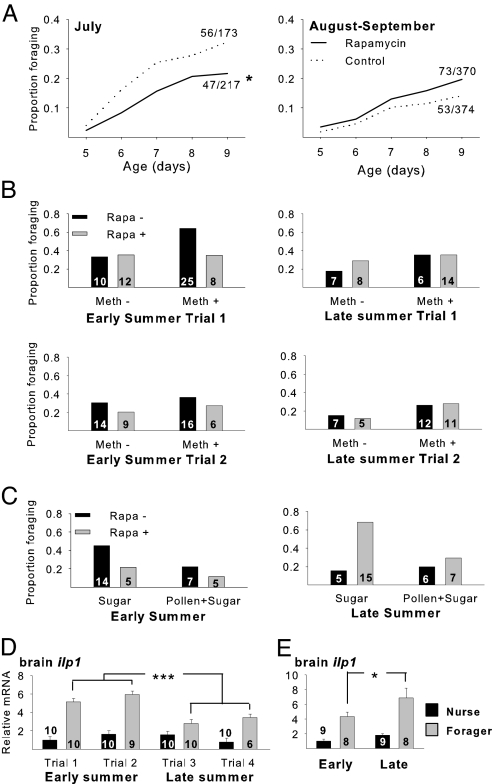
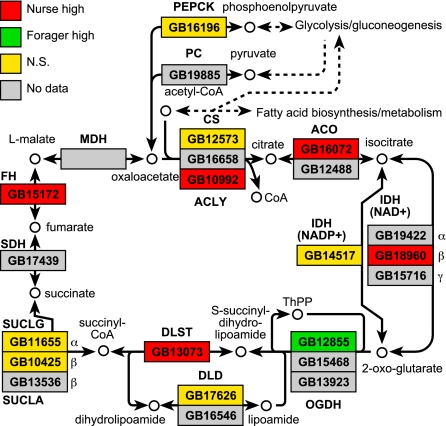
It has been proposed that one route of behavioral evolution involves novel regulation of conserved genes. Age-related division of labor in honey bee colonies, a highly derived behavioral system, involves the performance of different feeding-related tasks by different groups of individuals. Older bees acquire the colony's food by foraging for nectar and pollen, and the younger "nurse" bees feed larvae processed foods. The transition from hive work to foraging has been shown to be socially regulated and associated both with decreases in abdominal lipid stores and with increases in brain expression of genes implicated in feeding behavior in Drosophila melanogaster. Here we show that division of labor is influenced by a canonical regulator of food intake and energy balance in solitary species, the insulin/insulin-like growth factor signaling (IIS) pathway. Foragers had higher levels of IIS gene expression in the brain and abdomen than did nurses, despite their low lipid stores. These differences are likely nutritionally mediated because manipulations that induced low lipid stores in young bees also up-regulated these genes. Changes in IIS also causally influenced the timing of behavioral maturation: inhibition of the insulin-related target of rapamycin pathway delayed the onset of foraging in a seasonally dependent manner. In addition, pathway analyses of microarray data revealed that nurses and foragers differ in brain energy metabolism gene expression, but the differences are opposite predictions based on their insulin-signaling status. These results suggest that changes in the regulation of the IIS pathway are associated with social behavior.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Robinson GE, Ben-Shahar Y. Social behavior and comparative genomics: New genes or new gene regulation? Genes Brain Behav. 2002;1:197–203. - PubMed
-
- Carroll SB, Grenier JK, Weatherbee SD. From DNA to Diversity: Molecular Genetics and the Evolution of Animal Design. Malden, MA: Blackwell Publishing; 2005. p. 234.
-
- Robinson GE, Grozinger CM, Whitfield CW. Sociogenomics: social life in molecular terms. Nat Rev Genet. 2005;6:257–270. - PubMed
-
- Toth AL, Robinson GE. Evo-devo and the evolution of social behavior. Trends Genet. 2007;23:334–341. - PubMed
-
- Winston ML. The Biology of the Honey Bee. Cambridge, MA: Harvard Univ Press; 1987. p. 294.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
