

Identification and in vitro expansion of functional antigen-specific CD25+ FoxP3+ regulatory T cells in hepatitis C virus infection
- PMID: 18337568
- PMCID: PMC2346728
- DOI: 10.1128/JVI.01548-07
Identification and in vitro expansion of functional antigen-specific CD25+ FoxP3+ regulatory T cells in hepatitis C virus infection
Abstract
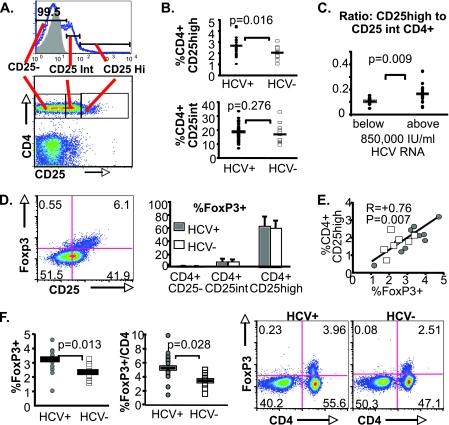
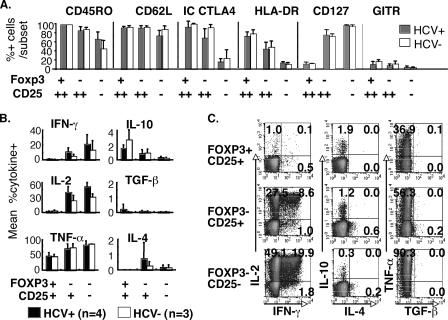
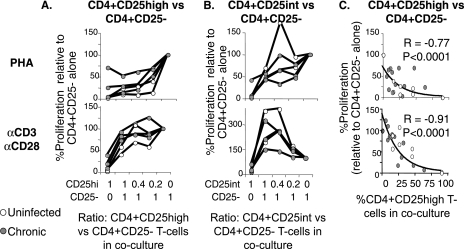
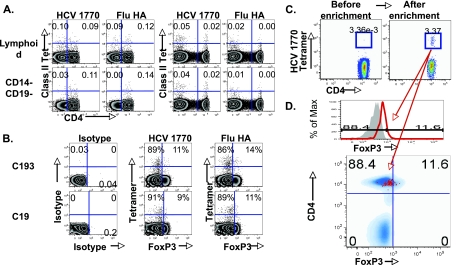
CD4(+)CD25(+) regulatory T cells (CD25(+) Tregs) play a key role in immune regulation. Since hepatitis C virus (HCV) persists with increased circulating CD4(+)CD25(+) T cells and virus-specific effector T-cell dysfunction, we asked if CD4(+)CD25(+) T cells in HCV-infected individuals are similar to natural Tregs in uninfected individuals and if they include HCV-specific Tregs using the specific Treg marker FoxP3 at the single-cell level. We report that HCV-infected patients display increased circulating FoxP3(+) Tregs that are phenotypically and functionally indistinguishable from FoxP3(+) Tregs in uninfected subjects. Furthermore, HCV-specific FoxP3(+) Tregs were detected in HCV-seropositive persons with antigen-specific expansion, major histocompatibility complex class II/peptide tetramer binding affinity, and preferential suppression of HCV-specific CD8 T cells. Transforming growth factor beta contributed to antigen-specific Treg expansion in vitro, suggesting that it may contribute to antigen-specific Treg expansion in vivo. Interestingly, FoxP3 expression was also detected in influenza virus-specific CD4 T cells. In conclusion, functionally active and virus-specific FoxP3(+) Tregs are induced in HCV infection, thus providing targeted immune regulation in vivo. Detection of FoxP3 expression in non-HCV-specific CD4 T cells suggests that immune regulation through antigen-specific Treg induction extends beyond HCV.
Figures



References
-
- Akbar, A. N., M. Vukmanovic-Stejic, L. S. Taams, and D. C. Macallan. 2007. The dynamic co-evolution of memory and regulatory CD4+ T cells in the periphery. Nat. Rev. Immunol. 7231-237. - PubMed
-
- Allan, S. E., S. Q. Crome, N. K. Crellin, L. Passerini, T. S. Steiner, R. Bacchetta, M. G. Roncarolo, and M. K. Levings. 2007. Activation-induced FOXP3 in human T effector cells does not suppress proliferation or cytokine production. Int. Immunol. 19345-354. - PubMed
-
- Baecher-Allan, C., J. A. Brown, G. J. Freeman, and D. A. Hafler. 2001. CD4+CD25high regulatory cells in human peripheral blood. J. Immunol. 1671245-1253. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
