

Role of nonstructural protein NS2A in flavivirus assembly
- PMID: 18337583
- PMCID: PMC2346727
- DOI: 10.1128/JVI.00002-08
Role of nonstructural protein NS2A in flavivirus assembly
Abstract
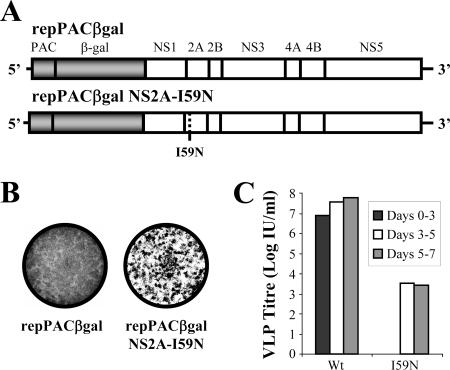
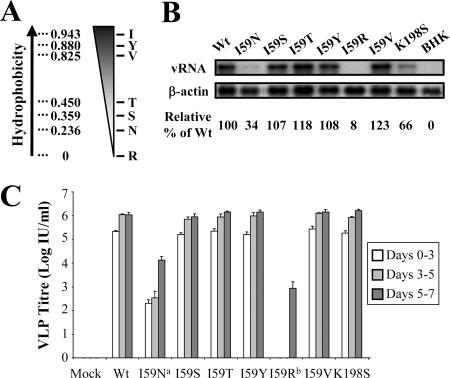
Flavivirus nonstructural (NS) proteins are involved in RNA replication and modulation of the host antiviral response; however, evidence is mounting that some NS proteins also have essential roles in virus assembly. Kunjin virus (KUN) NS2A is a small, hydrophobic, transmembrane protein that is part of the replication complex and inhibits interferon induction. Previously, we have shown that an isoleucine (I)-to-asparagine (N) substitution at position 59 of the NS2A protein blocked the production of secreted virus particles in cells electroporated with viral RNA carrying this mutation. We now show that prolonged incubation of mutant KUN NS2A-I59N replicon RNA, in an inducible BHK-derived packaging cell line (expressing KUN structural proteins C, prM, and E), generated escape mutants that rescued the secretion of infectious virus-like particles. Sequencing identified three groups of revertants that included (i) reversions to wild-type, hydrophobic Ile, (ii) pseudorevertants to more hydrophobic residues (Ser, Thr, and Tyr) at codon 59, and (iii) pseudorevertants retaining Asn at NS2A codon 59 but containing a compensatory mutation (Thr-to-Pro) at NS2A codon 149. Engineering hydrophobic residues at NS2A position 59 or the compensatory T149P mutation into NS2A-I59N replicon RNA restored the assembly of secreted virus-like particles in packaging cells. T149P mutation also rescued virus production when introduced into the full-length KUN RNA containing an NS2A-I59N mutation. Immunofluorescence and electron microscopy analyses of NS2A-I59N replicon-expressing cells showed a distinct lack of virus-induced membranes normally present in cells expressing wild-type replicon RNA. The compensatory mutation NS2A-T149P restored the induction of membrane structures to a level similar to those observed during wild-type replication. The results further confirm the role of NS2A in virus assembly, demonstrate the importance of hydrophobic residues at codon 59 in this process, implicate the involvement of NS2A in the biogenesis of virus-induced membranes, and suggest a vital role for the virus-induced membranes in virus assembly.
Figures



References
-
- Agirre, A., A. Barco, L. Carrasco, and J. L. Nieva. 2002. Viroporin-mediated membrane permeabilization. Pore formation by nonstructural poliovirus 2B protein. J. Biol. Chem. 27740434-40441. - PubMed
-
- Chambers, T. J., D. W. McCourt, and C. M. Rice. 1989. Yellow fever virus proteins NS2A, NS2B and NS4B: identification and partial N-terminal amino acid sequence analysis. Virology 169100-109. - PubMed
-
- Coia, G., M. D. Parker, G. Speight, M. E. Byrne, and E. G. Westaway. 1988. Nucleotide and complete amino acid sequences of Kunjin virus: definitive gene order and characteristics of the virus-specified proteins. J. Gen. Virol. 691-21. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
