

Structures of rhodopsin kinase in different ligand states reveal key elements involved in G protein-coupled receptor kinase activation
- PMID: 18339619
- PMCID: PMC2376226
- DOI: 10.1074/jbc.M708974200
Structures of rhodopsin kinase in different ligand states reveal key elements involved in G protein-coupled receptor kinase activation
Abstract
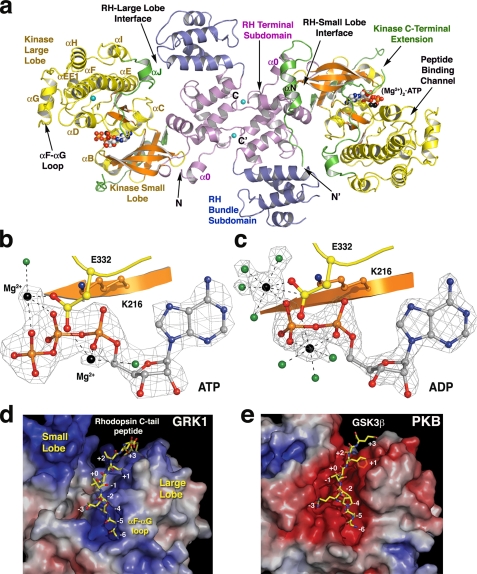
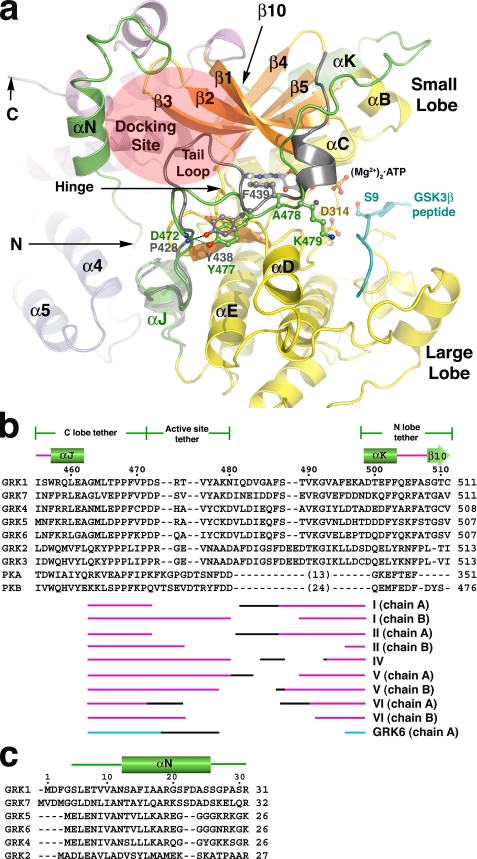
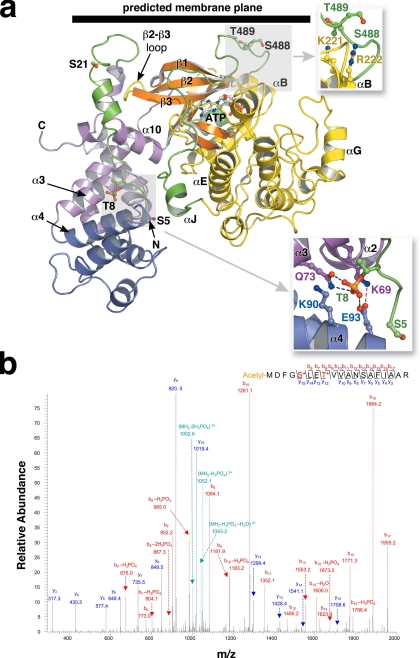
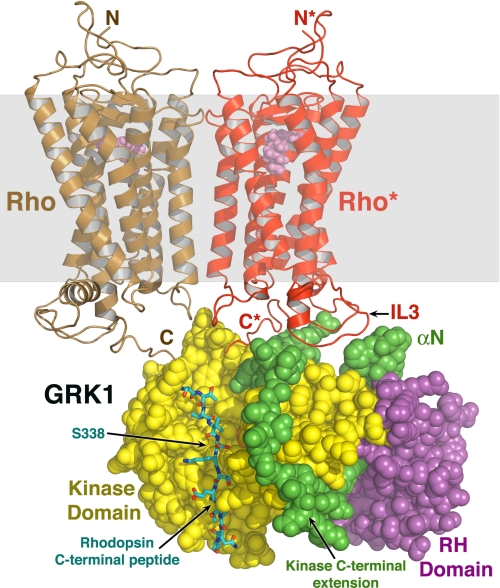
G protein-coupled receptor (GPCR) kinases (GRKs) phosphorylate activated heptahelical receptors, leading to their uncoupling from G proteins. Here we report six crystal structures of rhodopsin kinase (GRK1), revealing not only three distinct nucleotide-binding states of a GRK but also two key structural elements believed to be involved in the recognition of activated GPCRs. The first is the C-terminal extension of the kinase domain, which was observed in all nucleotide-bound GRK1 structures. The second is residues 5-30 of the N terminus, observed in one of the GRK1.(Mg2+)2.ATP structures. The N terminus was also clearly phosphorylated, leading to the identification of two novel phosphorylation sites by mass spectral analysis. Co-localization of the N terminus and the C-terminal extension near the hinge of the kinase domain suggests that activated GPCRs stimulate kinase activity by binding to this region to facilitate full closure of the kinase domain.
Figures
References
-
- Maeda, T., Imanishi, Y., and Palczewski, K. (2003) Prog. Retin Eye Res. 22 417-434 - PubMed
-
- Arshavsky, V. Y., Lamb, T. D., and Pugh, E. N., Jr. (2002) Annu. Rev. Physiol. 64 153-187 - PubMed
-
- Doan, T., Mendez, A., Detwiler, P. B., Chen, J., and Rieke, F. (2006) Science 313 530-533 - PubMed
-
- Yamamoto, S., Sippel, K. C., Berson, E. L., and Dryja, T. P. (1997) Nat. Genet. 15 175-178 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
