

Genomic evolution of the placenta using co-option and duplication and divergence
- PMID: 18340042
- PMCID: PMC2336813
- DOI: 10.1101/gr.071407.107
Genomic evolution of the placenta using co-option and duplication and divergence
Abstract
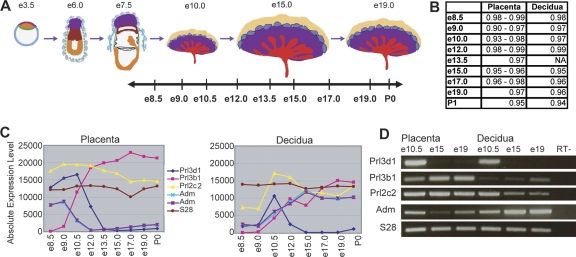
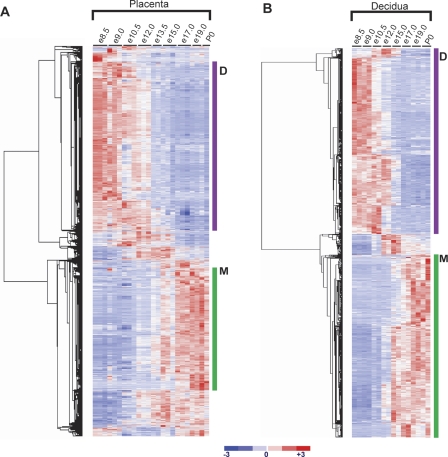
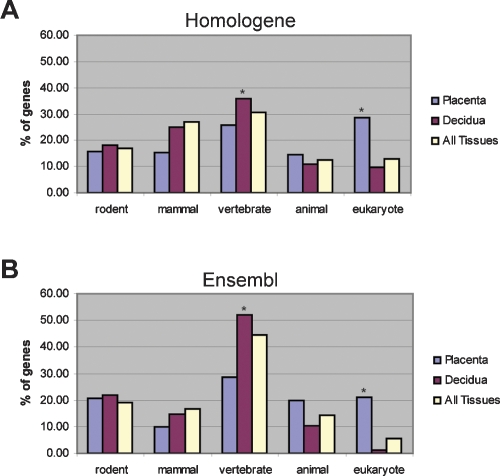
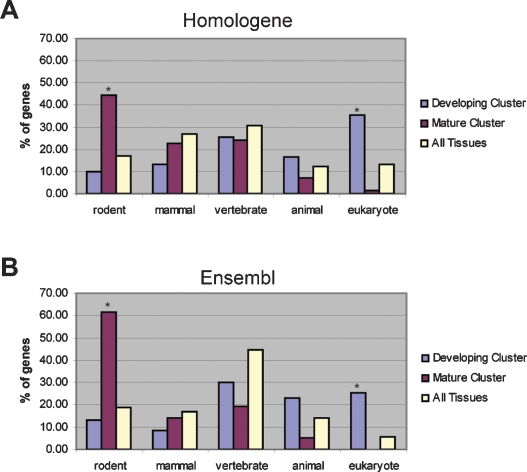
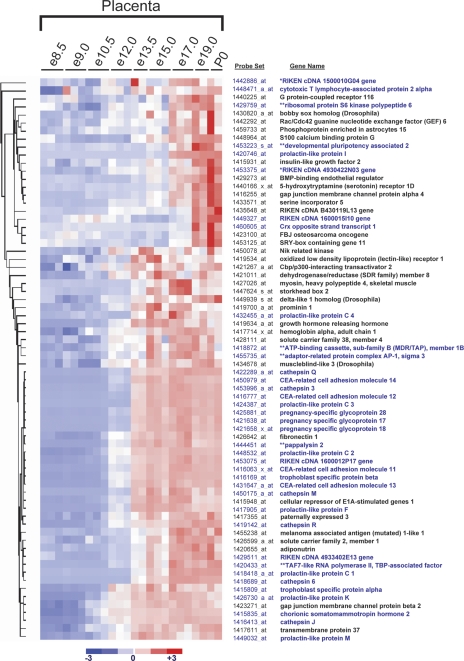
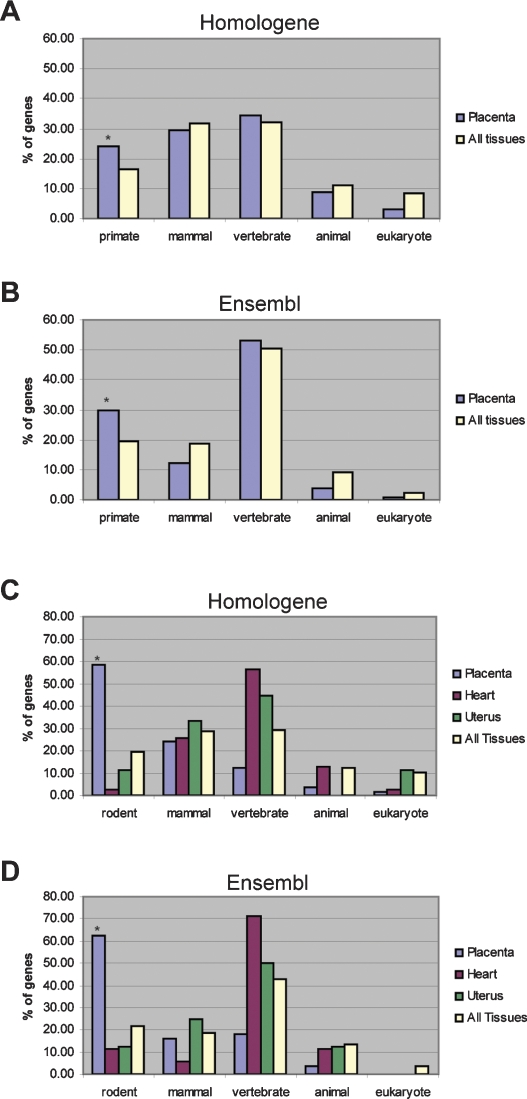
The invention of the placenta facilitated the evolution of mammals. How the placenta evolved from the simple structure observed in birds and reptiles into the complex organ that sustains human life is one of the great mysteries of evolution. By using a timecourse microarray analysis including the entire lifetime of the placenta, we uncover molecular and genomic changes that underlie placentation and find that two distinct evolutionary mechanisms were utilized during placental evolution in mice and human. Ancient genes involved in growth and metabolism were co-opted for use during early embryogenesis, likely enabling the accelerated development of extraembryonic tissues. Recently duplicated genes are utilized at later stages of placentation to meet the metabolic needs of a diverse range of pregnancy physiologies. Together, these mechanisms served to develop the specialized placenta, a novel structure that led to expansion of the eutherian mammal, including humankind.
Figures
References
-
- Ain R., Dai G., Dunmore J.H., Godwin A.R., Soares M.J., Dai G., Dunmore J.H., Godwin A.R., Soares M.J., Dunmore J.H., Godwin A.R., Soares M.J., Godwin A.R., Soares M.J., Soares M.J. A prolactin family paralog regulates reproductive adaptations to a physiological stressor. Proc. Natl. Acad. Sci. 2004;101:16543–16548. - PMC - PubMed
-
- Alam S.M., Ain R., Konno T., Ho-Chen J.K., Soares M.J., Ain R., Konno T., Ho-Chen J.K., Soares M.J., Konno T., Ho-Chen J.K., Soares M.J., Ho-Chen J.K., Soares M.J., Soares M.J. The rat prolactin gene family locus: Species-specific gene family expansion. Mamm. Genome. 2006;17:858–877. - PubMed
-
- Beauchemin N., Draber P., Dveksler G., Gold P., Gray-Owen S., Grunert F., Hammarstrom S., Holmes K.V., Karlsson A., Kuroki M., Draber P., Dveksler G., Gold P., Gray-Owen S., Grunert F., Hammarstrom S., Holmes K.V., Karlsson A., Kuroki M., Dveksler G., Gold P., Gray-Owen S., Grunert F., Hammarstrom S., Holmes K.V., Karlsson A., Kuroki M., Gold P., Gray-Owen S., Grunert F., Hammarstrom S., Holmes K.V., Karlsson A., Kuroki M., Gray-Owen S., Grunert F., Hammarstrom S., Holmes K.V., Karlsson A., Kuroki M., Grunert F., Hammarstrom S., Holmes K.V., Karlsson A., Kuroki M., Hammarstrom S., Holmes K.V., Karlsson A., Kuroki M., Holmes K.V., Karlsson A., Kuroki M., Karlsson A., Kuroki M., Kuroki M., et al. Redefined nomenclature for members of the carcinoembryonic antigen family. Exp. Cell Res. 1999;252:243–249. - PubMed
-
- Carter A.M., Mess A., Mess A. Evolution of the placenta in eutherian mammals. Placenta. 2006;28:259–262. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases