

Anionic phospholipids affect the rate and extent of flux through the mechanosensitive channel of large conductance MscL
- PMID: 18341289
- PMCID: PMC2566799
- DOI: 10.1021/bi702409t
Anionic phospholipids affect the rate and extent of flux through the mechanosensitive channel of large conductance MscL
Abstract
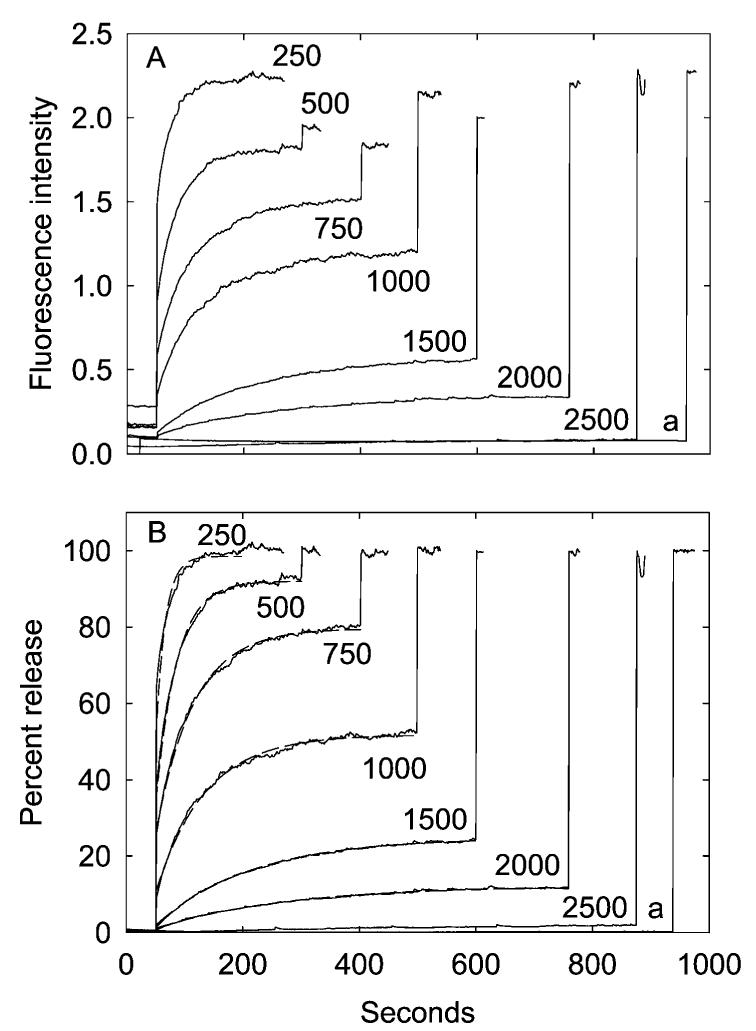
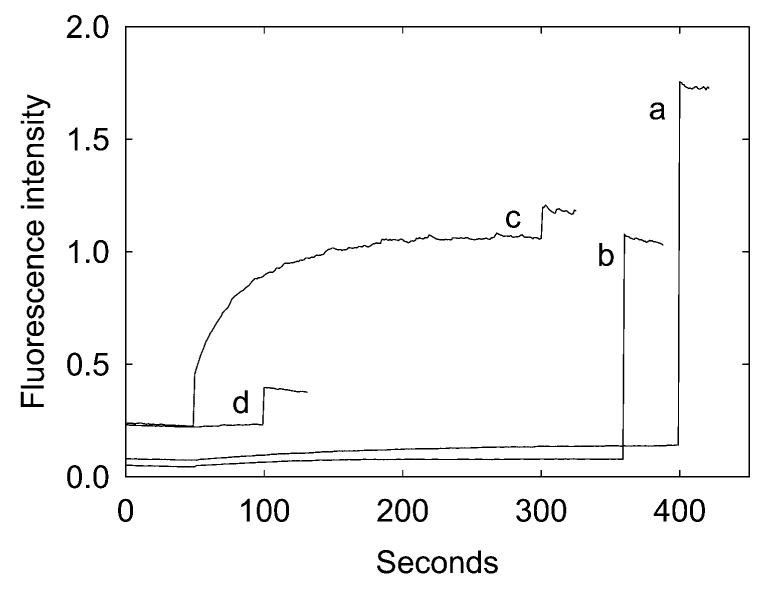
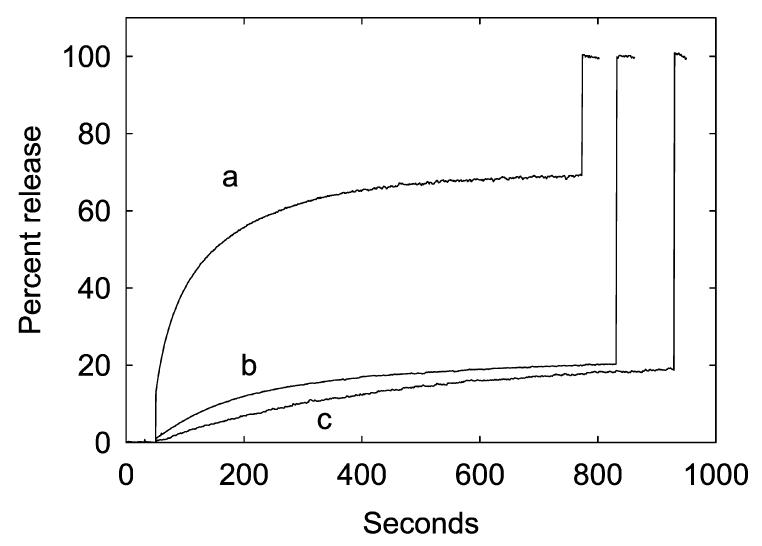
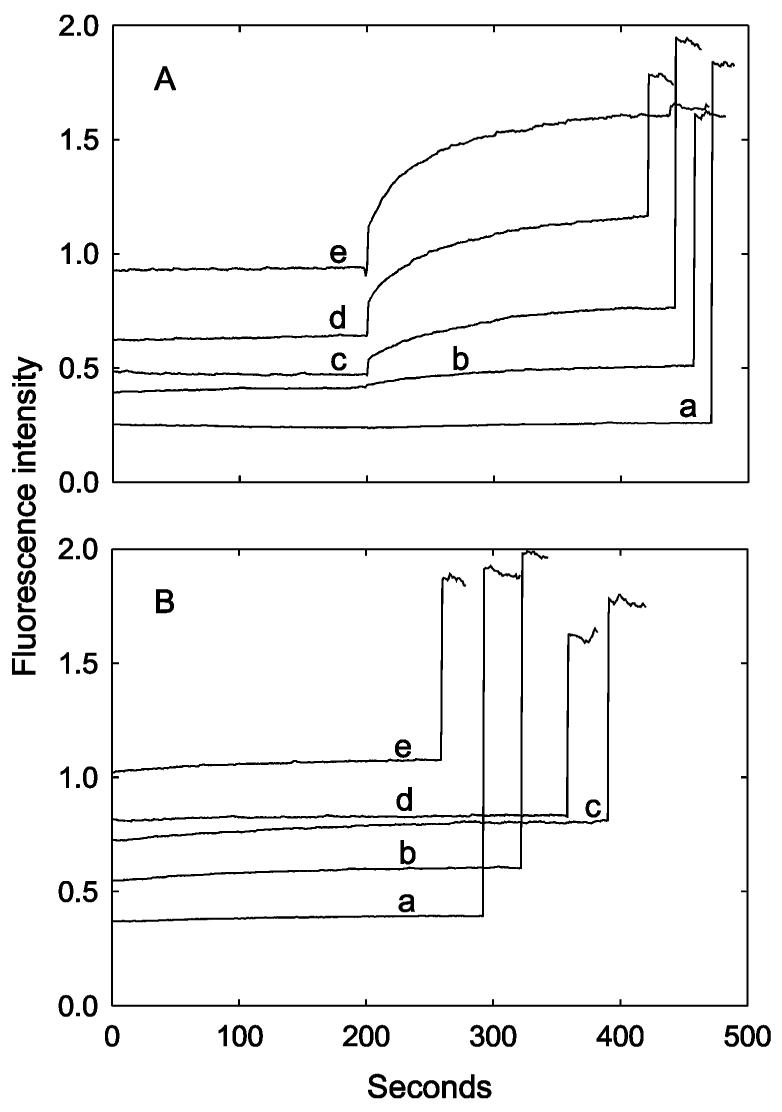
The mechanosensitive channel of large conductance MscL from Escherichia coli has been reconstituted into sealed vesicles, and the effects of lipid structure on the flux of the fluorescent molecule calcein through the open channel have been studied. The channel was opened by reaction of the G22C mutant of MscL with the reagent [2-(triethylammonium)ethyl]methanethiosulfonate (MTSET) which introduces five positive charges within the pore constriction. Flux through the channel was small when the lipid was phosphatidylcholine, but addition of the anionic lipids phosphatidylglycerol, phosphatidic acid, or cardiolipin up to 50 mol % resulted in increases in the amplitudes and rates of release of calcein. Similar effects were seen when either wild-type MscL or the G22C mutant was opened by osmotic pressure difference; rates of release of calcein were very slow in the absence of anionic lipid but increased with increasing concentrations of phosphatidylglycerol to 50 mol %. The observed partial release of trapped calcein following activation of MscL was attributed to the formation of a long-lived subconductance state of MscL following channel opening. Effects of anionic lipid were attributed to an increase in the rate of the transition from closed to fully open state and to a decrease in the rate of the transition from the fully open state to the subconductance state. Higher concentrations of anionic lipid led to a decrease in the rate and amplitude of release of calcein, possibly due to a decreased rate of flux through the open channel. In mixtures with anionic lipids, phosphatidylethanolamine resulted in lower rates and amplitude of release than phosphatidylcholine.
Figures








References
-
- Hamill OP, Martinac B. Molecular basis of mechanotransduction in living cells. Physiol. Rev. 2001;81:685–740. - PubMed
-
- Blount P, Iscla I, Moe PC, Li Y. MscL: the bacterial mechanosensitive channel of large conductance. Curr. Top. Membr. 2007;58:201–233.
-
- Lee AG. Lipid-protein interactions in biological membranes: a structural perspective. Biochim. Biophys. Acta. 2003;1612:1–40. - PubMed
-
- Lee AG. Lipid interactions with ion channels. Future Lipidol. 2006;1:103–113.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
