

doi: 10.1016/j.virol.2008.02.019.
Epub 2008 Mar 14.
Role of endocytosis and cathepsin-mediated activation in Nipah virus entry
Affiliations
- PMID: 18342904
- PMCID: PMC7103400
- DOI: 10.1016/j.virol.2008.02.019
Item in Clipboard
Role of endocytosis and cathepsin-mediated activation in Nipah virus entry
Virology.
.
Abstract
The recent discovery that the Nipah virus (NiV) fusion protein (F) is activated by endosomal cathepsin L raised the question if NiV utilize pH- and protease-dependent mechanisms of entry. We show here that the NiV receptor ephrin B2, virus-like particles and infectious NiV are internalized from the cell surface. However, endocytosis, acidic pH and cathepsin-mediated cleavage are not necessary for the initiation of infection of new host cells. Our data clearly demonstrate that proteolytic activation of the NiV F protein is required before incorporation into budding virions but not after virus entry.
Figures
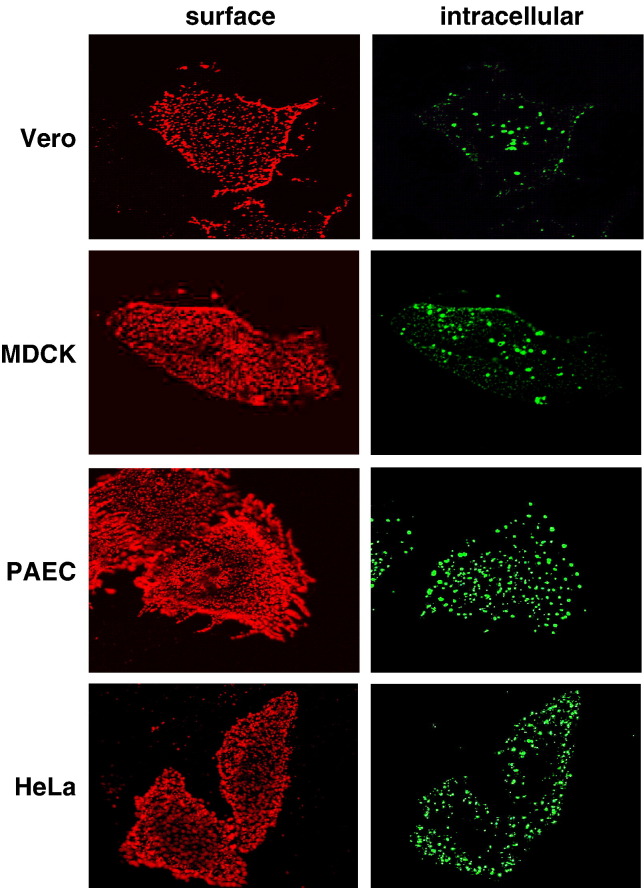
Endocytosis of EB2 in different cell lines. EB2-expressing cells (Vero, MDCK, PAEC, and HeLa) were incubated with EphB4/Fc for 1 h at 37 °C to allow binding and endocytosis to proceed. Surface-remained EphB4/Fc was stained with a rhodamine-conjugated anti-human IgG antibody (surface). After fixation and permeabilization, internalized EphB4/Fc was detected by a FITC-conjugated secondary antibody (intracellular).
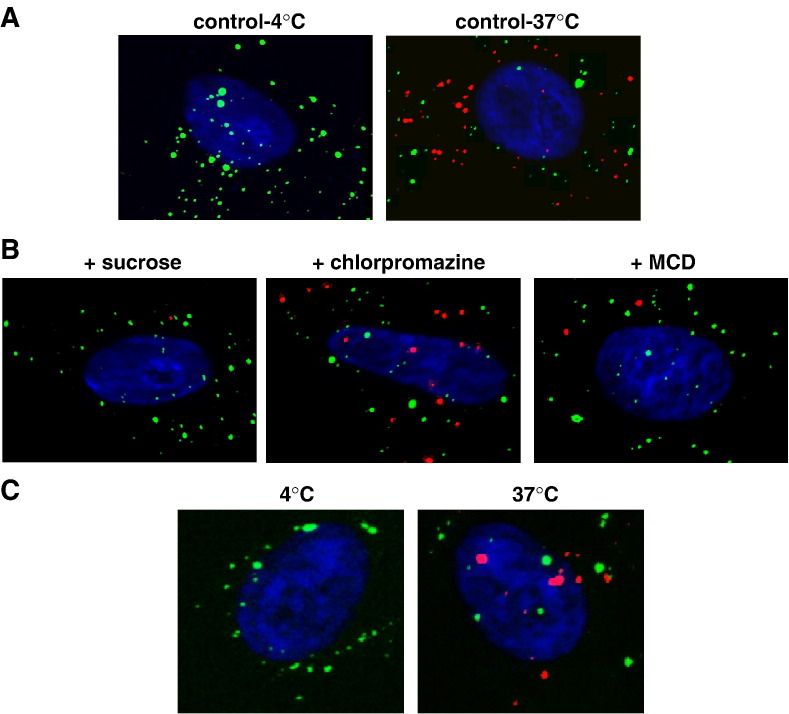
VLP- and virus-uptake assays. (A) Vero cells were incubated for 1 h at 4 °C (control-4 °C) or at 37 °C (control-37 °C) with purified NiV Gtag-containing VLPs in combination with a monoclonal antibody directed against the HA-tag. Surface-bound VLPs were visualized by incubation with FITC-conjugated anti-mouse IgG antibodies at 4 °C. After permeabilization with methanol–acetone, intracellular VLPs were stained with a rhodamine-conjugated secondary antibody. Nuclei were visualized by DAPI staining. Merged pictures of the DAPI, FITC and rhodamine fluorescence channels are shown. (B) Cells were preincubated for 30 min at 37 °C with the following endocytosis inhibitors: 0.45 M sucrose (sucrose), 25 µM chlorpromazine (chlorpromazine) or 5 mM β-methyl-cyclodextrin (MCD). Incubation with VLPs and HA-tag antibody was performed for 1 h at 37 °C in the presence or absence of the inhibitors. Surface-bound and intracellular VLPs were stained as described above. (C) Vero cells were incubated with infectious NiV particles for 2 h at 4 °C. After incubation with anti-NiV serum for 30 min on ice, cells were either shifted to 37 °C or kept at 4 °C for 20 min. Surface-bound and intracellular antibodies were visualized as described above.
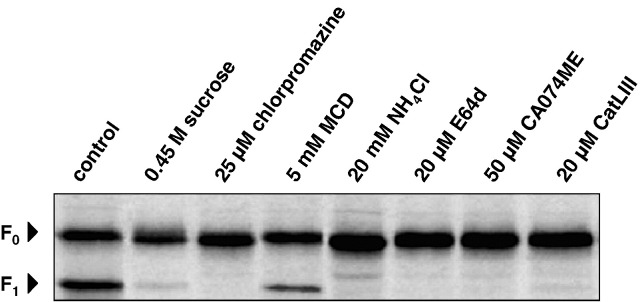
Effect of endocytosis inhibitors, NH4Cl, and cathepsin inhibitors on F protein cleavage. Vero cells expressing the NiV F protein were radiolabeled with [35S]-Promix for 10 min and were then incubated in chase medium containing the indicated inhibitors for 2 h. F proteins were immunoprecipitated from cell lysates, separated on a 12% SDS gel under reducing conditions, and subjected to autoradiography.
Effect of inhibitor treatments during and after virus entry on NiV infection. (A) Inhibitor treatment before and during virus entry. Vero cells were infected with NiV at a MOI of 0.2. Cells were treated with the indicated inhibitors for 1 h prior infection and for 1 h at 37 °C during virus adsorption and entry, or were left untreated (control). Then, inhibitors and input virus were removed, cells were washed and further incubated in medium for 24 h at 37 °C. After fixation and permeabilization, cells were incubated for 1 h on ice with a polyclonal anti-NiV serum and primary antibodies were detected with rhodamine-conjugated secondary antibodies. (B) Inhibitor treatment after virus entry. After infection of Vero cells with NiV for 1 h at 37 °C, cells were washed and then incubated in the absence (control) or presence of the indicated inhibitors. At 24 h p.i., immunostaining was performed as described above. There was no generalized toxicity effect of any of the inhibitors at the concentrations used in this experiment.
Effect of sucrose and NH4Cl treatment on influenza virus entry. Vero cells were infected with FPV at a MOI of 0.01. Cells were either left untreated (control) or incubated with the indicated inhibitors for 1 h prior infection and for 1 h during virus adsorption and entry. After washings to remove the inhibitors and input virus, residual virus attached to the cell surface was inactivated by acidic pH treatment. At 16 h p.i., cells were fixed and stained with a FPV-specific monoclonal antibody. Primary antibodies were detected with rhodamine-conjugated secondary antibodies and nuclei were visualized by DAPI staining.
F protein cleavage in purified NiV. Mock-sample and purified NiV were subjected to SDS-PAGE under reducing conditions and analyzed by Western blotting. Viral proteins were either detected with a polyclonal anti-NiV serum (anti-NiV) or with NiV F-specific antibodies (anti-F).
Similar articles
-
Activation of the Nipah virus fusion protein in MDCK cells is mediated by cathepsin B within the endosome-recycling compartment.J Virol. 2012 Apr;86(7):3736-45. doi: 10.1128/JVI.06628-11. Epub 2012 Jan 25. J Virol. 2012. PMID: 22278224 Free PMC article.
-
Ephrin-B2 expression critically influences Nipah virus infection independent of its cytoplasmic tail.Virol J. 2008 Dec 24;5:163. doi: 10.1186/1743-422X-5-163. Virol J. 2008. PMID: 19108727 Free PMC article.
-
Mutations in the G-H loop region of ephrin-B2 can enhance Nipah virus binding and infection.J Gen Virol. 2011 Sep;92(Pt 9):2142-2152. doi: 10.1099/vir.0.033787-0. Epub 2011 Jun 1. J Gen Virol. 2011. PMID: 21632558 Free PMC article.
-
Envelope-receptor interactions in Nipah virus pathobiology.Ann N Y Acad Sci. 2007 Apr;1102(1):51-65. doi: 10.1196/annals.1408.004. Ann N Y Acad Sci. 2007. PMID: 17470911 Free PMC article. Review.
-
Henipavirus membrane fusion and viral entry.Curr Top Microbiol Immunol. 2012;359:79-94. doi: 10.1007/82_2012_200. Curr Top Microbiol Immunol. 2012. PMID: 22427111 Review.
Cited by
-
Paramyxovirus Glycoproteins and the Membrane Fusion Process.Curr Clin Microbiol Rep. 2016 Sep;3(3):142-154. doi: 10.1007/s40588-016-0040-8. Epub 2016 Jul 5. Curr Clin Microbiol Rep. 2016. PMID: 28138419 Free PMC article.
-
Nipah virus fusion protein: Importance of the cytoplasmic tail for endosomal trafficking and bioactivity.Eur J Cell Biol. 2015 Jul-Sep;94(7-9):316-22. doi: 10.1016/j.ejcb.2015.05.005. Epub 2015 May 30. Eur J Cell Biol. 2015. PMID: 26059400 Free PMC article.
-
Characterization of the Bas-Congo virus glycoprotein and its function in pseudotyped viruses.J Virol. 2013 Sep;87(17):9558-68. doi: 10.1128/JVI.01183-13. Epub 2013 Jun 19. J Virol. 2013. PMID: 23785218 Free PMC article.
-
Host cell entry of respiratory syncytial virus involves macropinocytosis followed by proteolytic activation of the F protein.PLoS Pathog. 2013;9(4):e1003309. doi: 10.1371/journal.ppat.1003309. Epub 2013 Apr 11. PLoS Pathog. 2013. PMID: 23593008 Free PMC article.
-
Selective inhibitor of endosomal trafficking pathways exploited by multiple toxins and viruses.Proc Natl Acad Sci U S A. 2013 Dec 10;110(50):E4904-12. doi: 10.1073/pnas.1302334110. Epub 2013 Nov 4. Proc Natl Acad Sci U S A. 2013. PMID: 24191014 Free PMC article.
References
-
- Adams R.H. Vascular patterning by Eph receptor tyrosine kinases and ephrins. Semin. Cell Dev. Biol. 2002;13:55–60. - PubMed
-
- Cantin C., Holguera J., Ferreira L., Villar E., Munoz-Barroso I. Newcastle disease virus may enter cells by caveolae-mediated endocytosis. J. Gen. Virol. 2007;88:559–569. - PubMed
-
- Chen C., Vincent O., Jin J., Weisz O.A., Montelaro R.C. Functions of early (AP-2) and late (AIP1/ALIX) endocytic proteins in equine infectious anemia virus budding. J. Biol. Chem. 2005;280:40474–40480. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
