

Motivationally neutral stimulation of the nucleus basalis induces specific behavioral memory
- PMID: 18343695
- PMCID: PMC2496874
- DOI: 10.1016/j.nlm.2008.02.001
Motivationally neutral stimulation of the nucleus basalis induces specific behavioral memory
Abstract
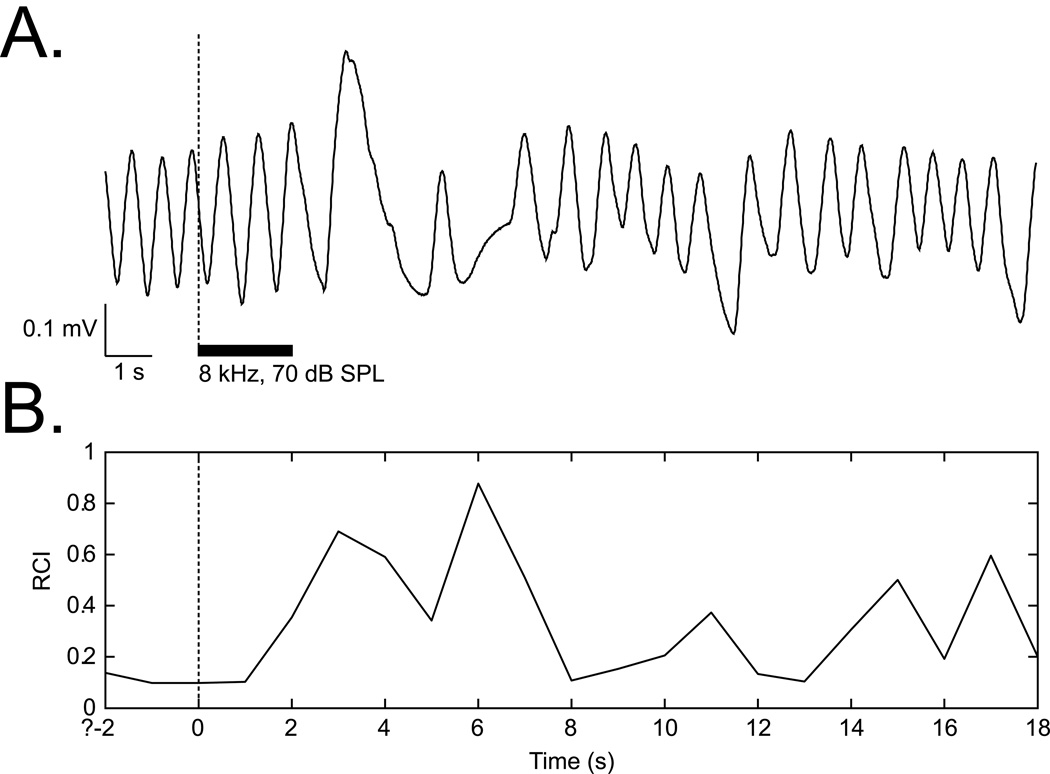
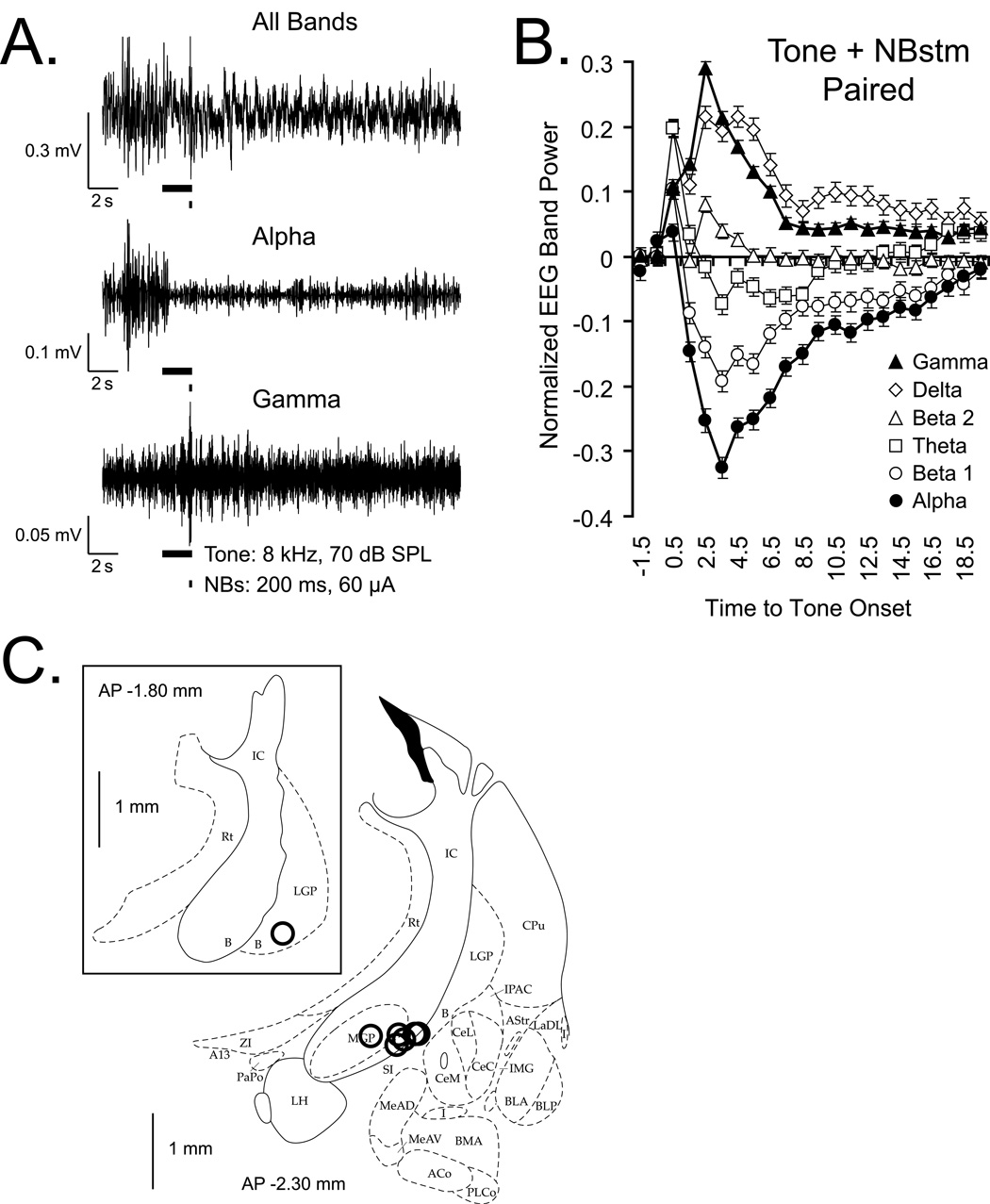
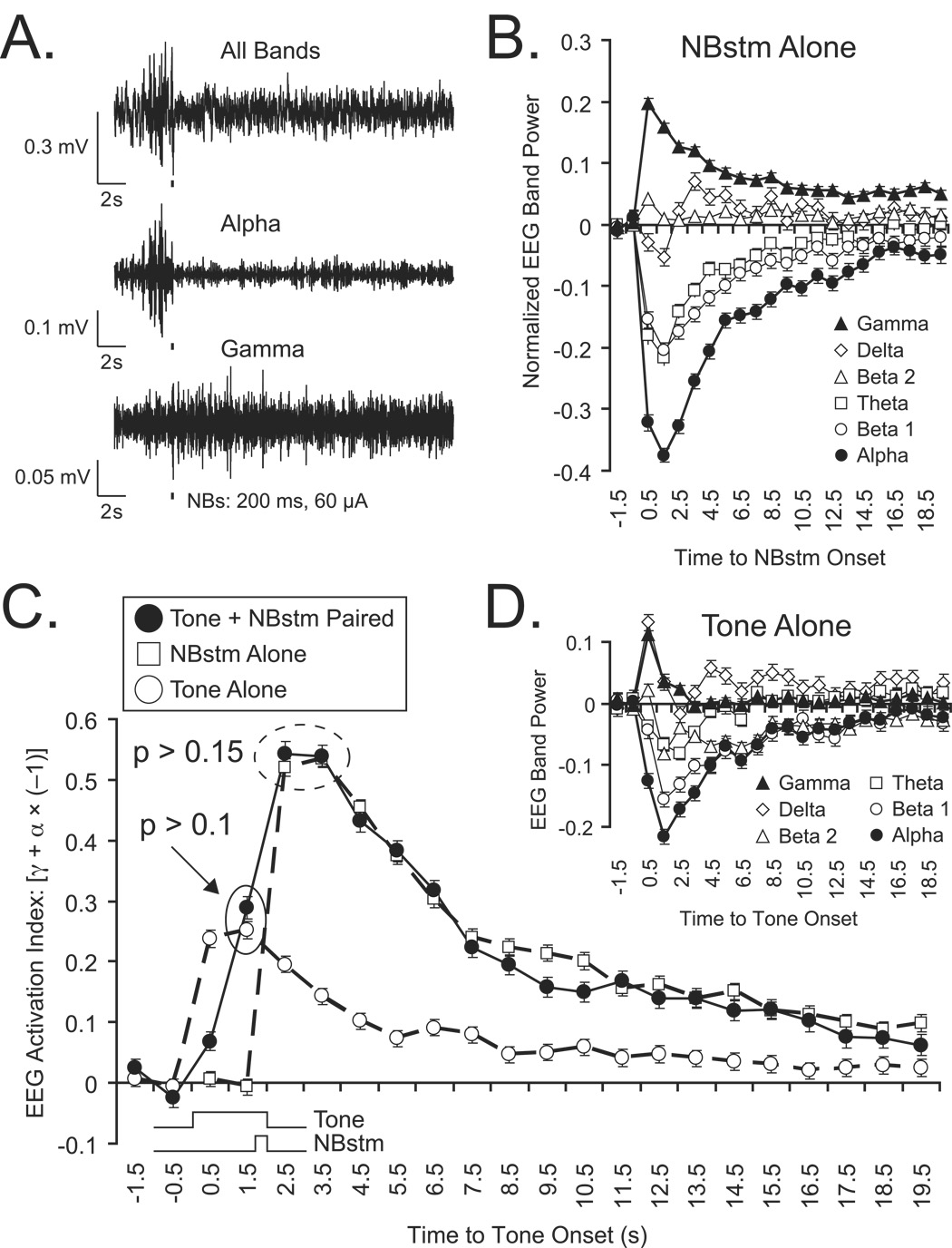
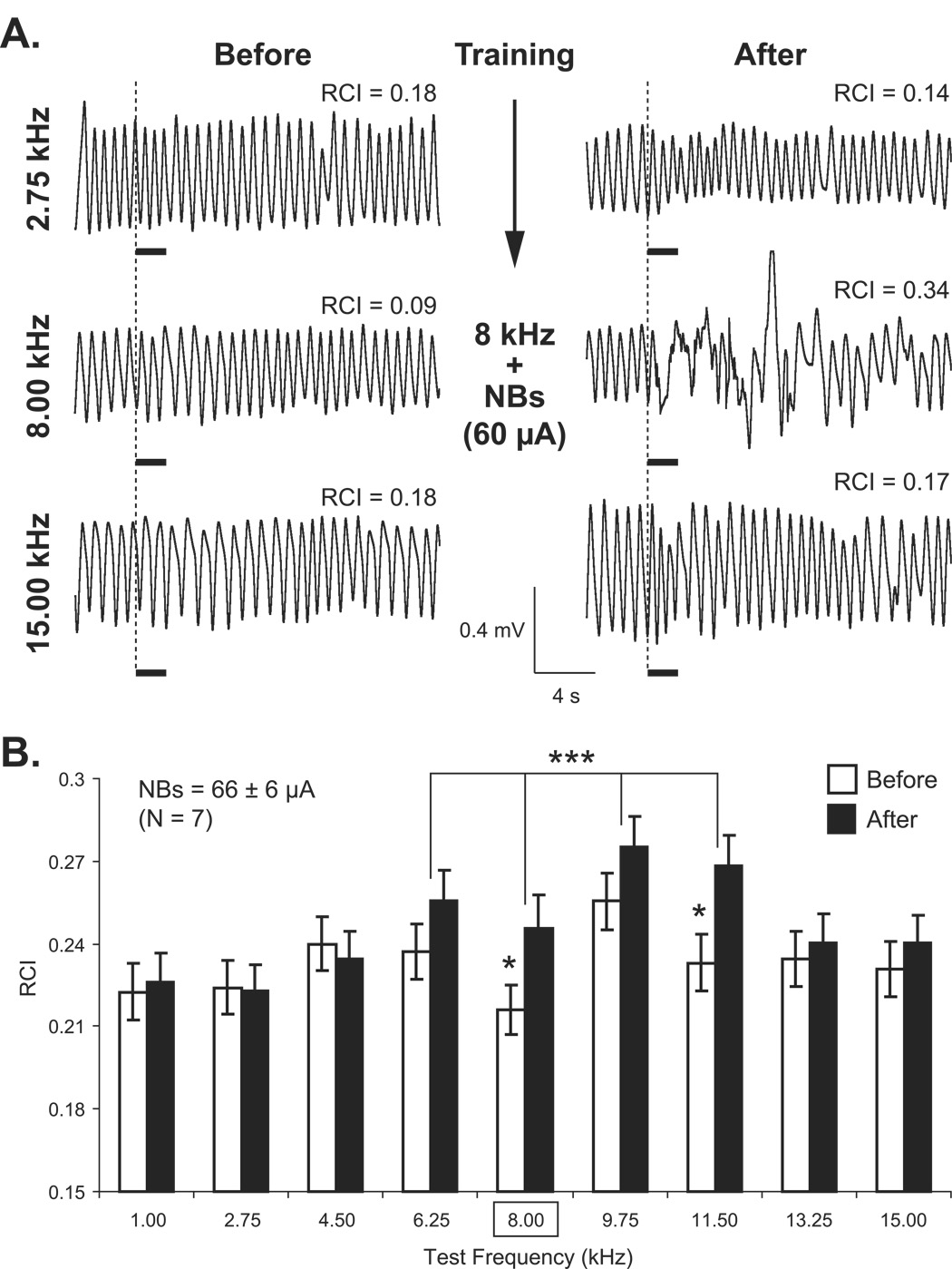
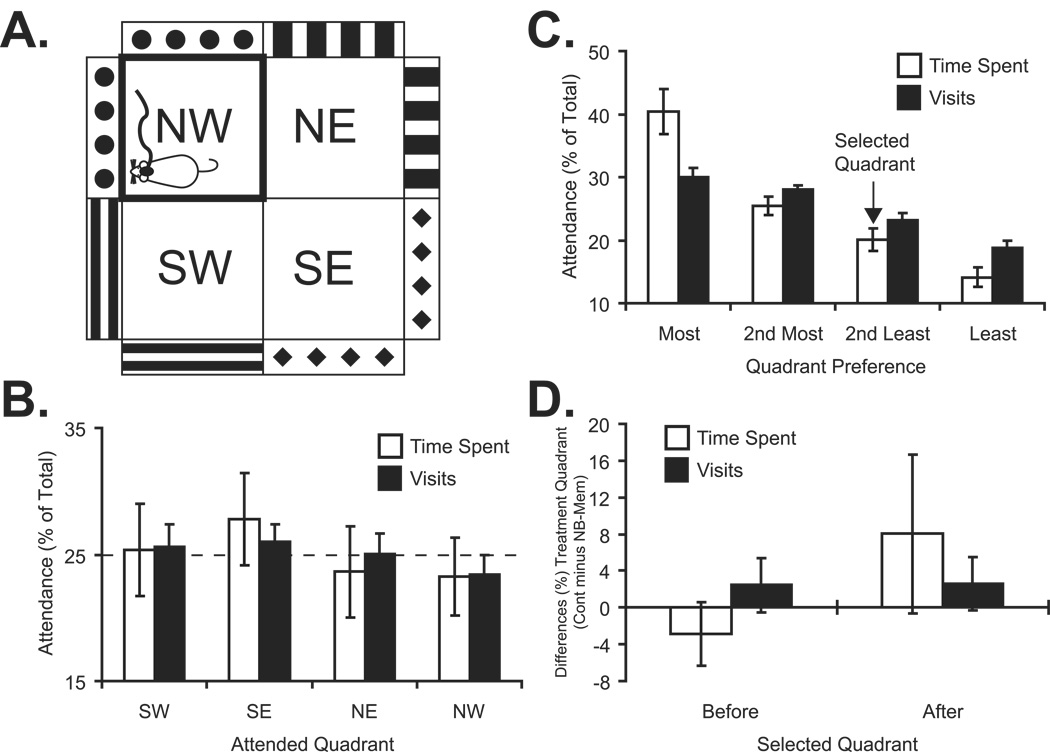
The cholinergic system has been implicated in learning and memory. The nucleus basalis (NB) provides acetylcholine (ACh) to the cerebral cortex. Pairing a tone with NB stimulation (NBstm) to alter cortical state induces both associative specific tuning plasticity in the primary auditory cortex (A1) and associative specific auditory behavioral memory. NB-induced memory has major features of natural memory that is induced by pairing a tone with motivational reinforcers, e.g., food or shock, suggesting that the cholinergic system may be a "final common pathway" whose activation promotes memory storage. Alternatively, NB stimulation might itself be motivationally significant, either rewarding or punishing. To investigate these alternatives, adult male rats (n=7) first formed a specific NB-induced memory (CS=8.0kHz, 2.0s paired with NBstm, ISI=1.8s, 200 trials), validated by post-training (24h) frequency generalization gradients (1-15kHz) of respiration interruption that were specific to the CS frequency. Thereafter, they received the same level of NBstm that had induced memory, while confined to one quadrant of an arena, and later tested for place-preference, i.e., avoidance or seeking of the quadrant of NBstm. This NBstm group exhibited neither preference for nor against the stimulated quadrant, compared to sham-operated subjects (n=7). The findings indicate that specific associative memory can be induced by direct activation of the NB without detectable motivational effects of NB stimulation. These results are concordant with a memory-promoting role for the nucleus basalis that places it "downstream" of motivational systems, which activate it to initiate the storage of the current state of its cholinergic targets.
Figures
Similar articles
-
Behavioral memory induced by stimulation of the nucleus basalis: effects of contingency reversal.Neurobiol Learn Mem. 2009 Mar;91(3):298-309. doi: 10.1016/j.nlm.2008.12.010. Epub 2009 Jan 23. Neurobiol Learn Mem. 2009. PMID: 19168141 Free PMC article.
-
Rapid induction of specific associative behavioral memory by stimulation of the nucleus basalis in the rat.Neurobiol Learn Mem. 2006 Jul;86(1):47-65. doi: 10.1016/j.nlm.2005.12.010. Epub 2006 Feb 8. Neurobiol Learn Mem. 2006. PMID: 16466937 Free PMC article.
-
Induction of behavioral associative memory by stimulation of the nucleus basalis.Proc Natl Acad Sci U S A. 2002 Mar 19;99(6):4002-7. doi: 10.1073/pnas.062057099. Proc Natl Acad Sci U S A. 2002. PMID: 11904444 Free PMC article.
-
The nucleus basalis and memory codes: auditory cortical plasticity and the induction of specific, associative behavioral memory.Neurobiol Learn Mem. 2003 Nov;80(3):268-84. doi: 10.1016/s1074-7427(03)00072-8. Neurobiol Learn Mem. 2003. PMID: 14521869 Review.
-
Auditory associative memory and representational plasticity in the primary auditory cortex.Hear Res. 2007 Jul;229(1-2):54-68. doi: 10.1016/j.heares.2007.01.004. Epub 2007 Jan 17. Hear Res. 2007. PMID: 17344002 Free PMC article. Review.
Cited by
-
Specific auditory memory induced by nucleus basalis stimulation depends on intrinsic acetylcholine.Neurobiol Learn Mem. 2008 Sep;90(2):443-54. doi: 10.1016/j.nlm.2008.05.010. Epub 2008 Jun 23. Neurobiol Learn Mem. 2008. PMID: 18573347 Free PMC article.
-
Behavioral memory induced by stimulation of the nucleus basalis: effects of contingency reversal.Neurobiol Learn Mem. 2009 Mar;91(3):298-309. doi: 10.1016/j.nlm.2008.12.010. Epub 2009 Jan 23. Neurobiol Learn Mem. 2009. PMID: 19168141 Free PMC article.
-
Short-Term Effects of Vagus Nerve Stimulation on Learning and Evoked Activity in Auditory Cortex.eNeuro. 2021 Jun 28;8(3):ENEURO.0522-20.2021. doi: 10.1523/ENEURO.0522-20.2021. Print 2021 May-Jun. eNeuro. 2021. PMID: 34088737 Free PMC article.
-
Optogenetic Dissection of the Basal Forebrain Neuromodulatory Control of Cortical Activation, Plasticity, and Cognition.J Neurosci. 2015 Oct 14;35(41):13896-903. doi: 10.1523/JNEUROSCI.2590-15.2015. J Neurosci. 2015. PMID: 26468190 Free PMC article. Review.
-
Gamma band plasticity in sensory cortex is a signature of the strongest memory rather than memory of the training stimulus.Neurobiol Learn Mem. 2013 Sep;104:49-63. doi: 10.1016/j.nlm.2013.05.001. Epub 2013 May 10. Neurobiol Learn Mem. 2013. PMID: 23669065 Free PMC article.
References
-
- Bao S, Chan VT, Merzenich MM. Cortical remodelling induced by activity of ventral tegmental dopamine neurons. Nature. 2001;412(6842):79–83. - PubMed
-
- Bardo MT, Rowlett JK, Harris MJ. Conditioned place preference using opiate and stimulant drugs: A meta-analysis. Neuroscience and Biobehavioral Reviews. 1995;19(1):39–51. - PubMed
-
- Bigl V, Woolf NJ, Butcher LL. Cholinergic projections from the basal forebrain to frontal, parietal, temporal, occipital, and cingulate cortices: A combined fluorescent tracer and acetylcholinesterase analysis. Brain Research Bulletin. 1982;8(6):727–749. - PubMed
-
- Bjordahl TS, Dimyan MA, Weinberger NM. Induction of long-term receptive field plasticity in the auditory cortex of the waking guinea pig by stimulation of the nucleus basalis. Behavioral Neuroscience. 1998;112(3):467–479. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
