

Changes in sensory-evoked synaptic activation of motoneurons after spinal cord injury in man
- PMID: 18344559
- PMCID: PMC2566952
- DOI: 10.1093/brain/awn050
Changes in sensory-evoked synaptic activation of motoneurons after spinal cord injury in man
Abstract
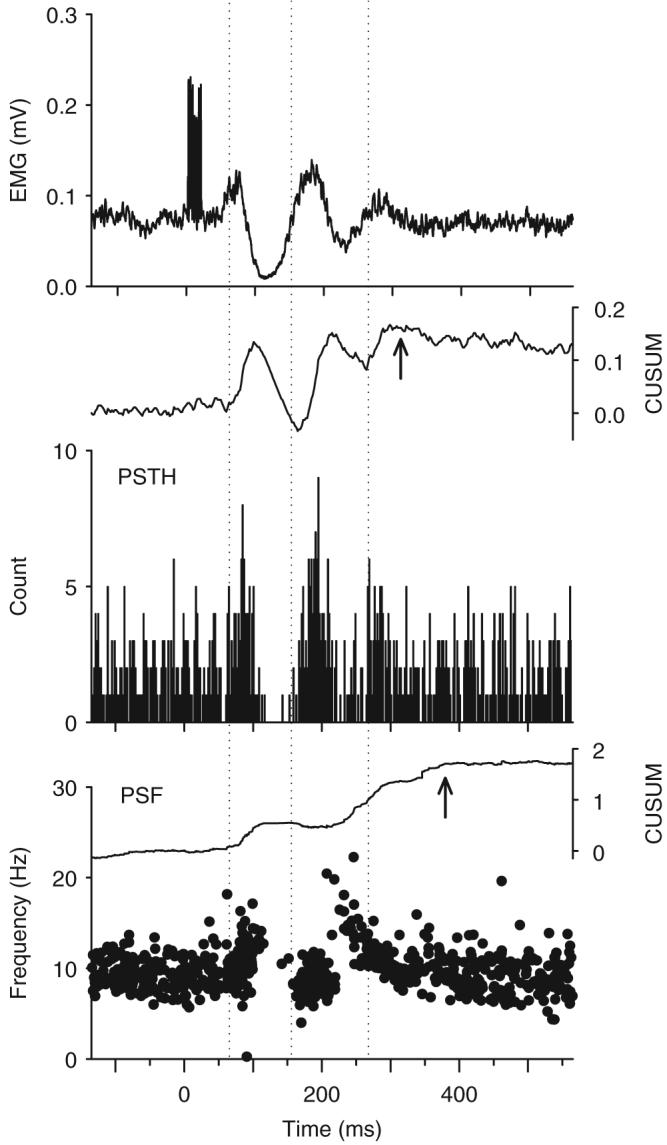
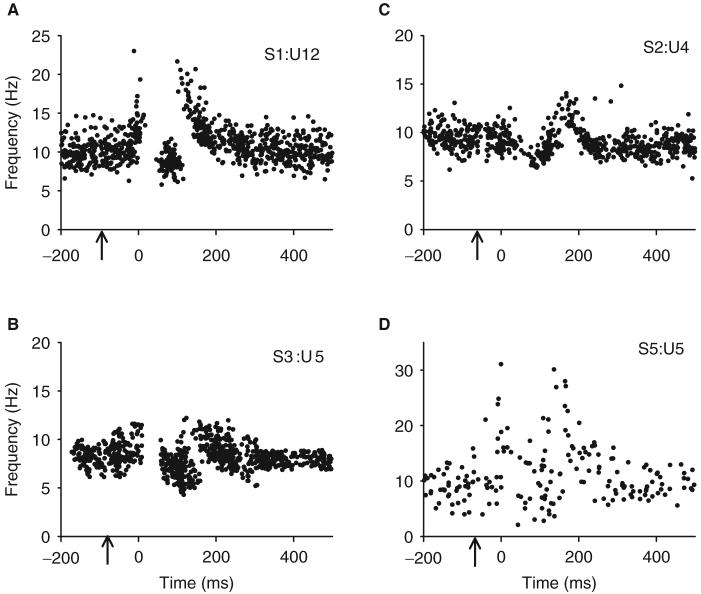
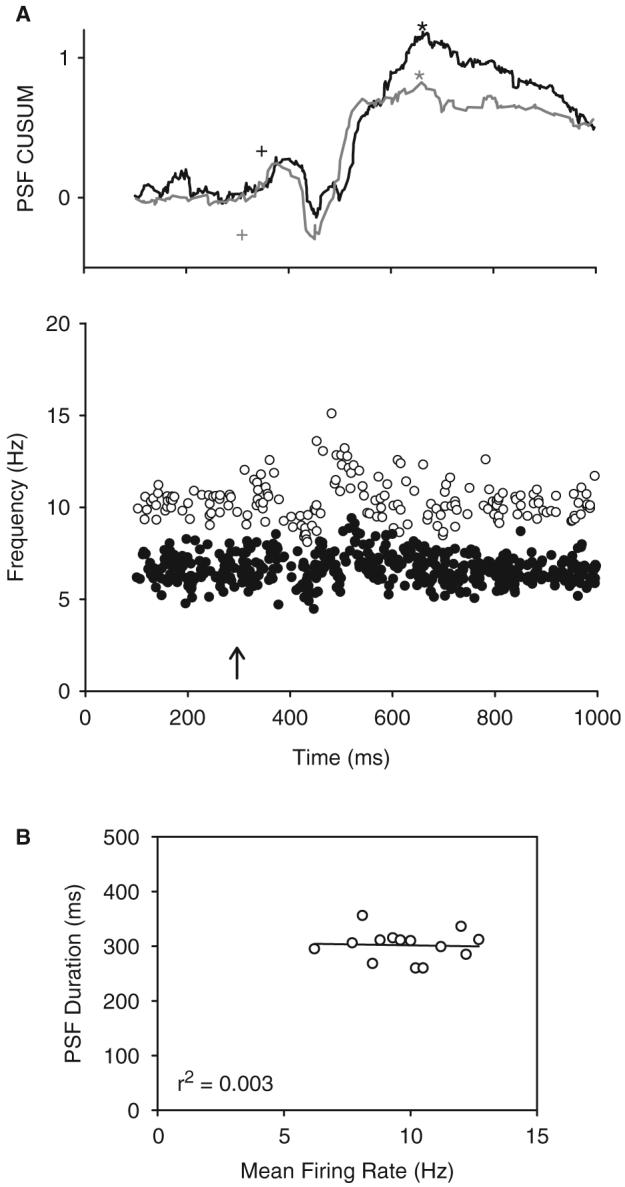
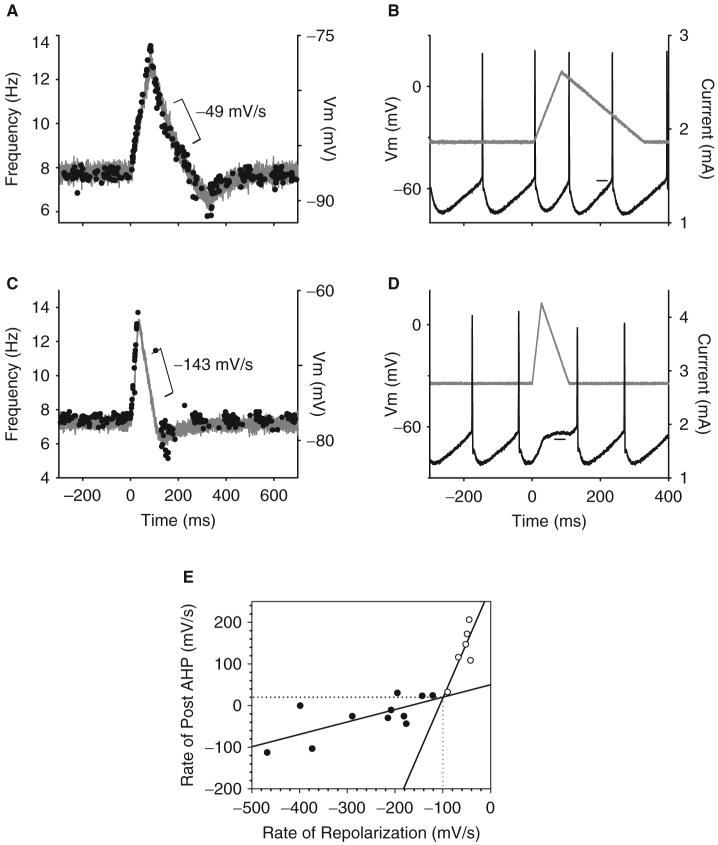
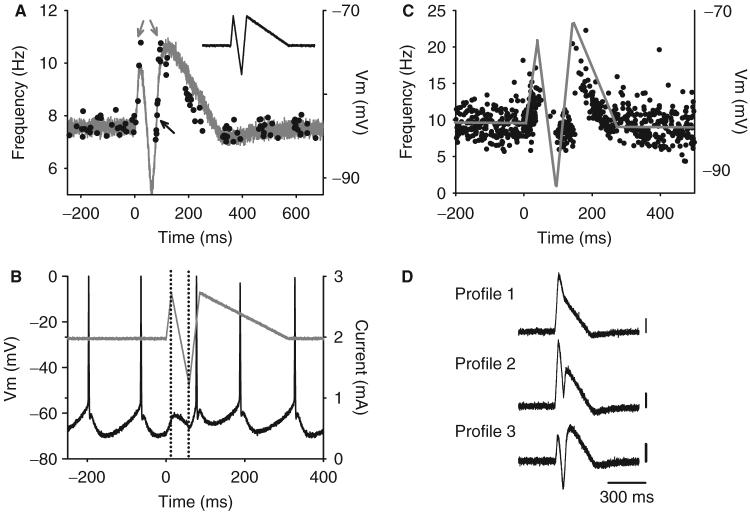
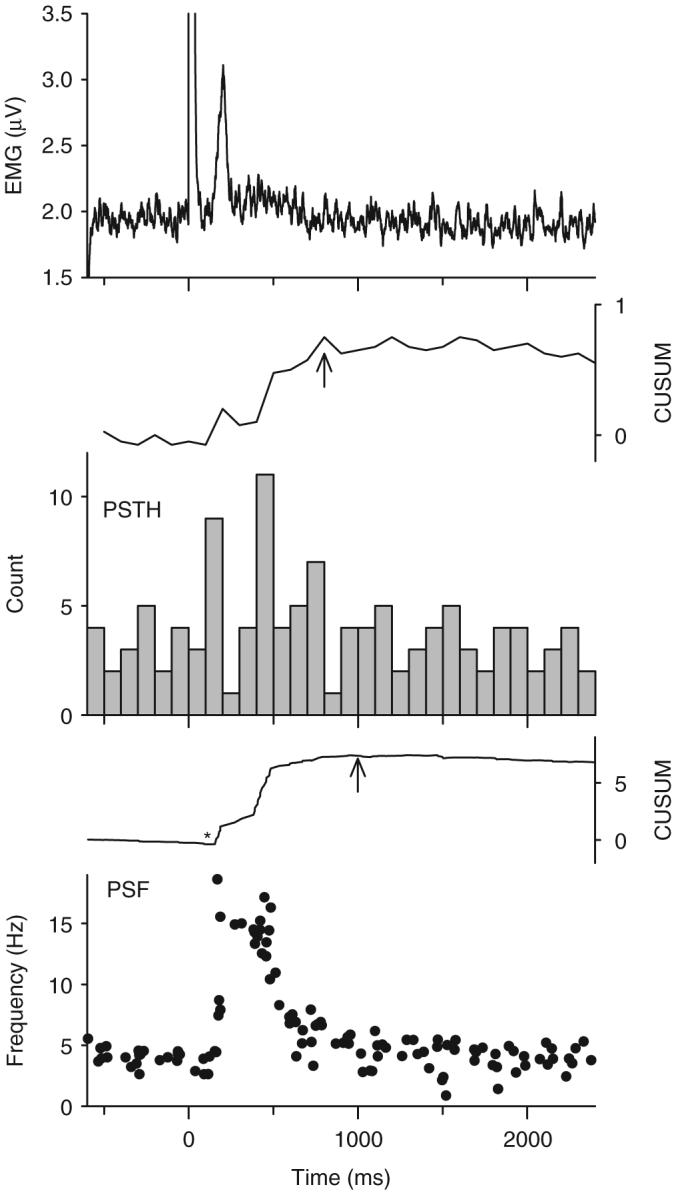
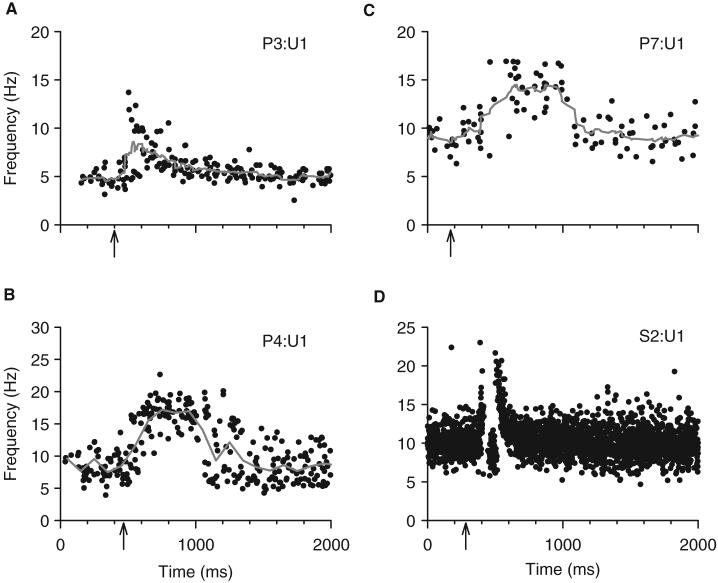
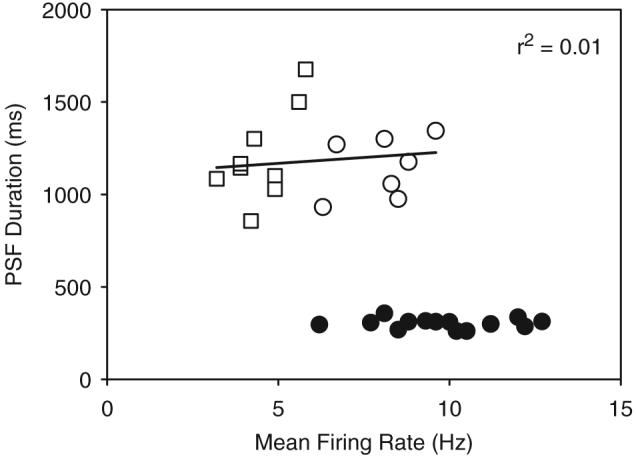
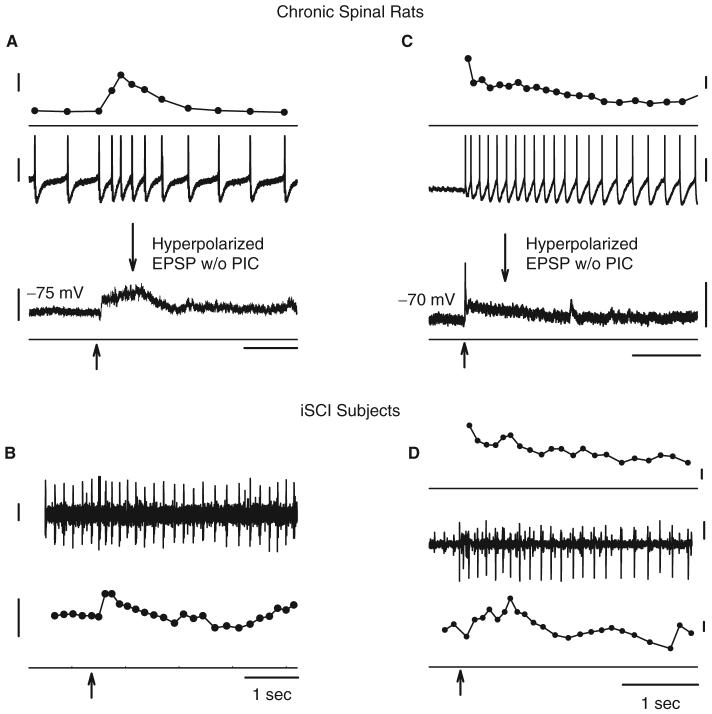
Following spinal cord injury (SCI), prolonged muscle spasms are readily triggered by brief sensory stimuli. Animal and indirect human studies have shown that a substantial portion of the depolarization of motoneurons during a muscle spasm comes from the activation of persistent inward currents (PICs). The brief (single pulse) sensory stimuli that trigger the PICs and muscle spasms in chronically spinalized animals evoke excitatory post-synaptic potentials (EPSPs) that are broadened to more than 500 ms, the duration of depolarization required to activate a PIC in the motoneuron. Thus, in humans, we investigated if post-synaptic potentials (PSPs) evoked from brief (<20 ms) sensory stimulation are changed after SCI and if they are broadened to > or =500 ms to more readily activate motoneuron PICs and muscle spasms. To estimate both the shape and duration of PSPs in human subjects we used peristimulus frequencygrams (PSFs), which are plots of the instantaneous firing frequency of tonically active single motor units that are time-locked to the occurrence of the sensory stimulus. PSFs in response to cutaneomuscular stimulation of the medial arch or toe of the foot, a sensory stimulus that readily triggers muscle spasms, were compared between non-injured control subjects and in spastic subjects with chronic (>1 year), incomplete SCI. In non-injured controls, a single shock or brief (<20 ms) train of cutaneomuscular stimulation produced PSFs consisting of a 300 ms increase in firing rate above baseline with an interposed period of reduced firing. Parallel intracellular experiments in motoneurons of adult rats revealed that a 300 ms EPSP with a fast intervening inhibitory PSP (IPSP) reproduced the PSF recorded in non-injured subjects. In contrast, the same brief sensory stimulation in subjects with chronic SCI produced PSFs of comparatively long duration (1200 ms) with no evidence for IPSP activation, as reflected by a lack of reduced firing rates after the onset of the PSF. Thus, unlike non-injured controls, the motoneurons of subjects with chronic SCI are activated by very long periods of pure depolarization from brief sensory activation. It is likely that these second-long EPSPs securely recruit slowly activating PICs in motoneurons that are known to mediate, in large part, the many seconds-long activation of motoneurons during involuntary muscle spasms.
Figures
References
-
- Baker LL, Chandler SH. Characterization of postsynaptic potentials evoked by sural nerve stimulation in hindlimb motoneurons from acute and chronic spinal cats. Brain Res. 1987;420:340–50. - PubMed
-
- Bennett DJ, Li Y, Sanelli L. Role of NMDA in spasticity following sacral spinal cord injury in rats. Soc Neurosci Abstr. 2001a;31:933.11.
-
- Bennett DJ, Li Y, Siu M. Plateau potentials in sacrocaudal motoneurons of chronic spinal rats, recorded in vitro. J Neurophysiol. 2001b;86:1955–71. - PubMed
-
- Bennett DJ, Sanelli L, Cooke CL, Harvey PJ, Gorassini MA. Spastic long-lasting reflexes in the awake rat after sacral spinal injury. J. Neurophysiol. 2004;91:2247–58. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
