

Wall teichoic acid deficiency in Staphylococcus aureus confers selective resistance to mammalian group IIA phospholipase A(2) and human beta-defensin 3
- PMID: 18347049
- PMCID: PMC2346714
- DOI: 10.1128/IAI.01705-07
Wall teichoic acid deficiency in Staphylococcus aureus confers selective resistance to mammalian group IIA phospholipase A(2) and human beta-defensin 3
Abstract
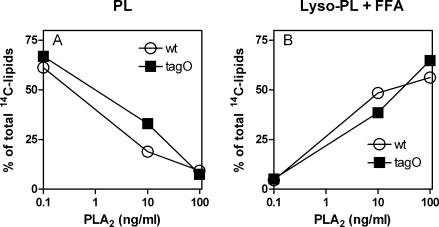
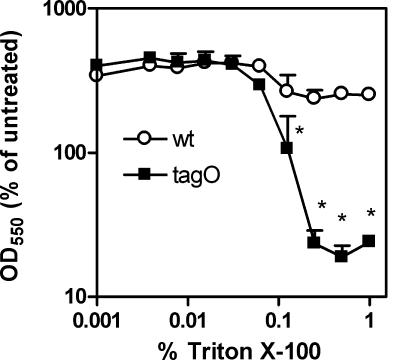
Wall teichoic acids (WTAs) and membrane lipoteichoic acids (LTAs) are the major polyanionic polymers in the envelope of Staphylococcus aureus. WTAs in S. aureus play an important role in bacteriophage attachment and bacterial adherence to certain host cells, suggesting that WTAs are exposed on the cell surface and could also provide necessary binding sites for cationic antimicrobial peptides and proteins (CAMPs). Highly cationic mammalian group IIA phospholipase A(2) (gIIA PLA(2)) kills S. aureus at nanomolar concentrations by an action(s) that depends on initial electrostatic interactions, cell wall penetration, membrane phospholipid (PL) degradation, and activation of autolysins. A tagO mutant of S. aureus that lacks WTA is up to 100-fold more resistant to PL degradation and killing by gIIA PLA(2) and CAMP human beta-defensin 3 (HBD-3) but has the sensitivity of the wild type (wt) to other CAMPs, such as Magainin II amide, hNP1-3, LL-37, and lactoferrin. In contrast, there is little or no difference in either gIIA PLA(2) activity toward cell wall-depleted protoplasts of the wt and tagO strains of S. aureus or in binding of gIIA PLA(2) to wt and tagO strains. Scanning and transmission electron microscopy reveal increased surface protrusions in the S. aureus tagO mutant that might account for reduced activity of bound gIIA PLA(2) and HBD-3 toward the tagO mutant. In summary, the absence of WTA in S. aureus causes a selective increase in bacterial resistance to gIIA PLA(2) and HBD-3, the former apparently by reducing access and/or activity of bound antibacterial enzyme to the bacterial membrane.
Figures




References
-
- Beers, S. A., A. G. Buckland, R. S. Koduri, W. Cho, M. H. Gelb, and D. C. Wilton. 2002. The antibacterial properties of secreted phospholipases A2: a major physiological role for the group IIA enzyme that depends on the very high pI of the enzyme to allow penetration of the bacterial cell wall. J. Biol. Chem. 2771788-1793. - PubMed
-
- Bligh, E. G., and W. J. Dyer. 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37911-917. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
