

Myosin V and Kinesin act as tethers to enhance each others' processivity
- PMID: 18347333
- PMCID: PMC2290781
- DOI: 10.1073/pnas.0711531105
Myosin V and Kinesin act as tethers to enhance each others' processivity
Abstract
Organelle transport to the periphery of the cell involves coordinated transport between the processive motors kinesin and myosin V. Long-range transport takes place on microtubule tracks, whereas final delivery involves shorter actin-based movements. The concept that motors only function on their appropriate track required further investigation with the recent observation that myosin V undergoes a diffusional search on microtubules. Here we show, using single-molecule techniques, that a functional consequence of myosin V's diffusion on microtubules is a significant enhancement of the processive run length of kinesin when both motors are present on the same cargo. The degree of run length enhancement correlated with the net positive charge in loop 2 of myosin V. On actin, myosin V also undergoes longer processive runs when kinesin is present on the same cargo. The process that causes run length enhancement on both cytoskeletal tracks is electrostatic. We propose that one motor acts as a tether for the other and prevents its diffusion away from the track, thus allowing more steps to be taken before dissociation. The resulting run length enhancement likely contributes to the successful delivery of cargo in the cell.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
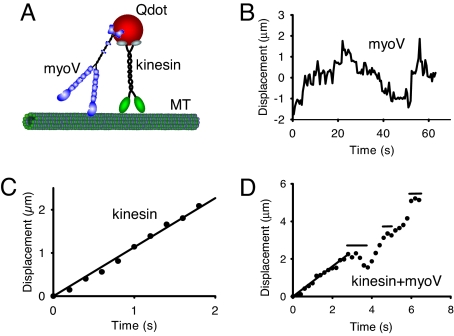
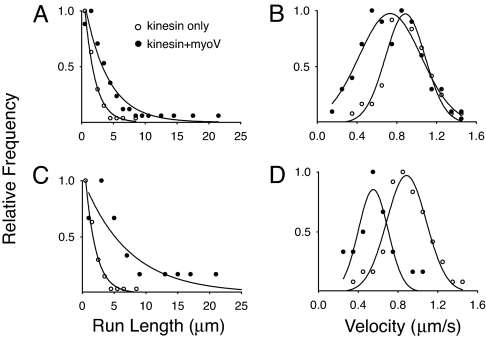
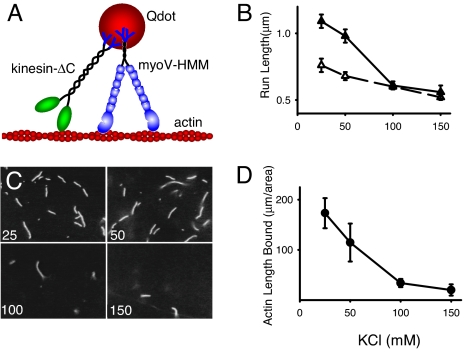
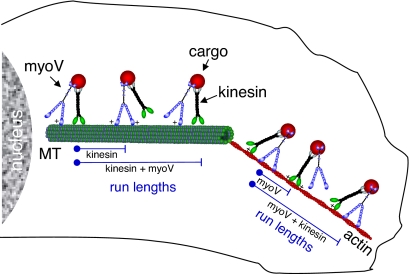
References
-
- Huang JD, Brady ST, Richards BW, Stenolen D, Resau JH, Copeland NG, Jenkins NA. Direct interaction of microtubule- and actin-based transport motors. Nature. 1999;397:267–270. - PubMed
-
- Hodges AR, Krementsova EB, Trybus KM. Engineering the processive run length of Myosin V. J Biol Chem. 2007;282:27192–27197. - PubMed
-
- Berezuk MA, Schroer TA. Dynactin enhances the processivity of kinesin-2. Traffic. 2007;8:124–129. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
