

Highly specific gene silencing by artificial miRNAs in rice
- PMID: 18350165
- PMCID: PMC2262943
- DOI: 10.1371/journal.pone.0001829
Highly specific gene silencing by artificial miRNAs in rice
Abstract
Background: Endogenous microRNAs (miRNAs) are potent negative regulators of gene expression in plants and animals. Artificial miRNAs (amiRNAs)-designed to target one or several genes of interest-provide a new and highly specific approach for effective post-transcriptional gene silencing (PTGS) in plants.
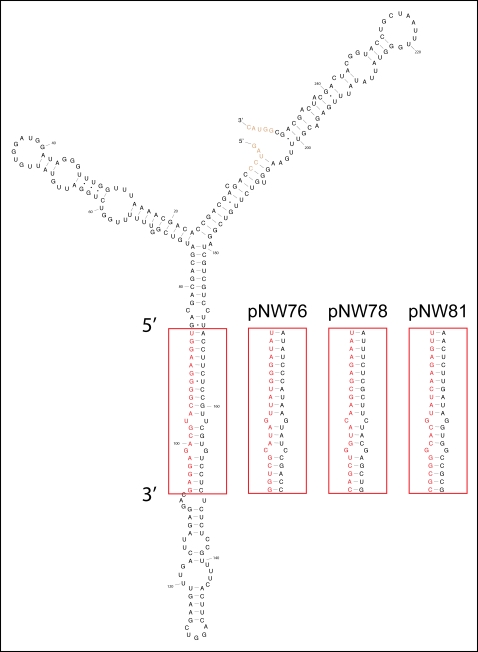
Methodology: We devised an amiRNA-based strategy for both japonica and indica type strains of cultivated rice, Oryza sativa. Using an endogenous rice miRNA precursor and customized 21mers, we designed amiRNA constructs targeting three different genes (Pds, Spl11, and Eui1/CYP714D1). Upon constitutive expression of these amiRNAs in the varieties Nipponbare (japonica) and IR64 (indica), the targeted genes are down-regulated by amiRNA-guided cleavage of the transcripts, resulting in the expected mutant phenotypes. The effects are highly specific to the target gene, the transgenes are stably inherited and they remain effective in the progeny.
Conclusion/significance: Our results not only show that amiRNAs can efficiently trigger gene silencing in a monocot crop, but also that amiRNAs can effectively modulate agronomically important traits in varieties used in modern breeding programs. We provide all software tools and a protocol for the design of rice amiRNA constructs, which can be easily adapted to other crops. The approach is suited for candidate gene validation, comparative functional genomics between different varieties, and for improvement of agronomic performance and nutritional value.
Conflict of interest statement
Figures



References
-
- Song XJ, Huang W, Shi M, Zhu MZ, Lin HX. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet. 2007;39:623–630. - PubMed
-
- Niu QW, Lin SS, Reyes JL, Chen KC, Wu HW, et al. Expression of artificial microRNAs in transgenic Arabidopsis thaliana confers virus resistance. Nat Biotechnol. 2006;24:1420–1428. - PubMed
-
- Sasaki A, Ashikari M, Ueguchi-Tanaka M, Itoh H, Nishimura A, et al. Green revolution: a mutant gibberellin-synthesis gene in rice. Nature. 2002;416:701–702. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
