

Structural elements regulating amyloidogenesis: a cholinesterase model system
- PMID: 18350169
- PMCID: PMC2265548
- DOI: 10.1371/journal.pone.0001834
Structural elements regulating amyloidogenesis: a cholinesterase model system
Abstract
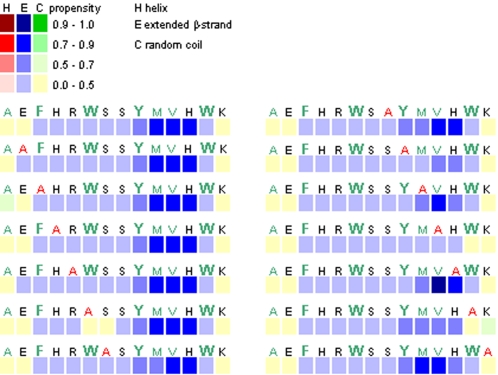
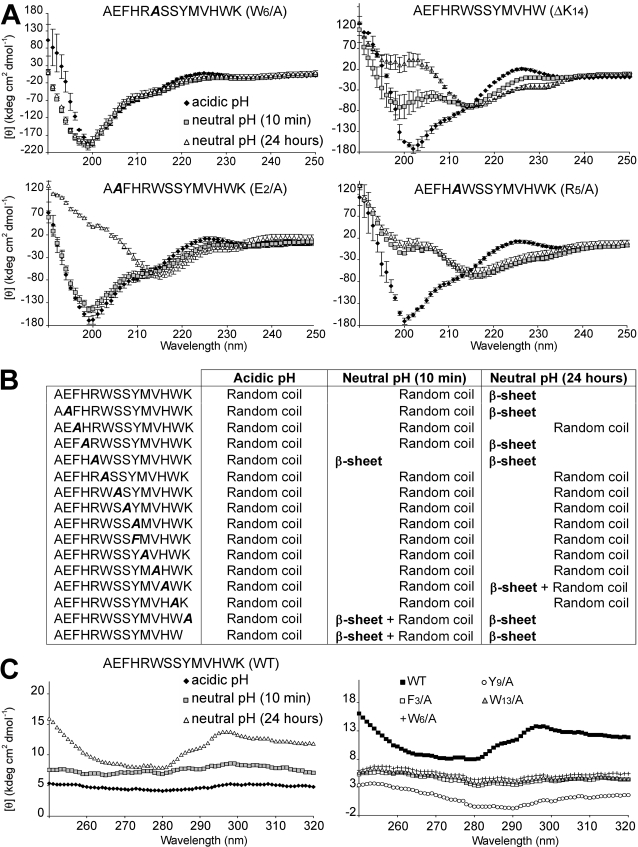
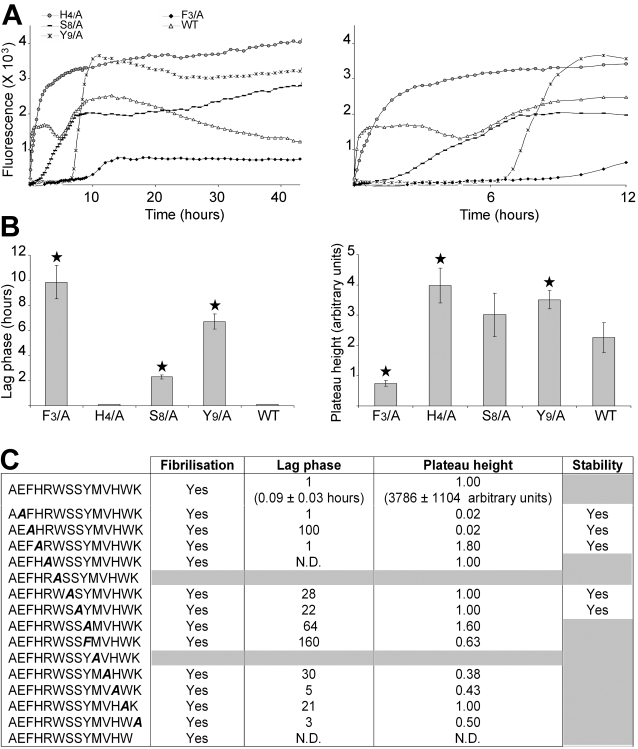
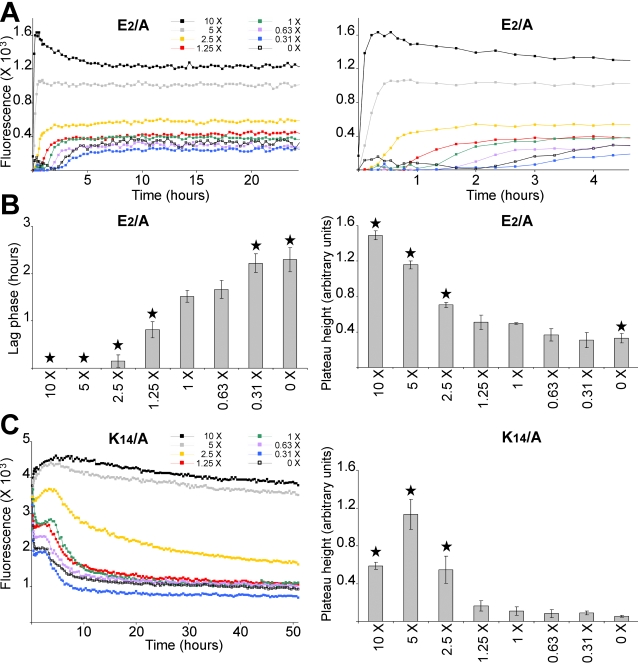
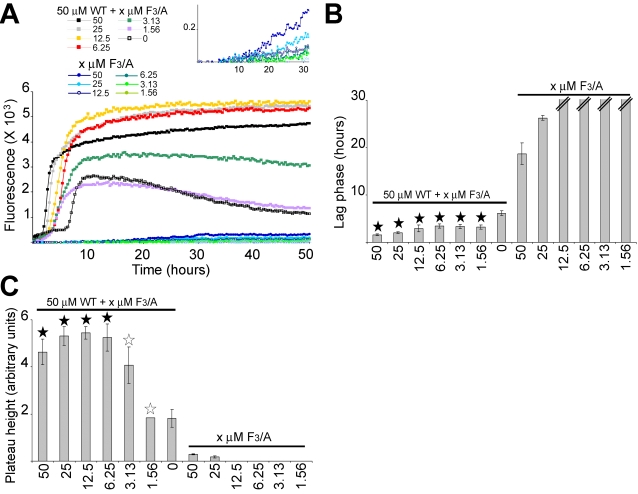
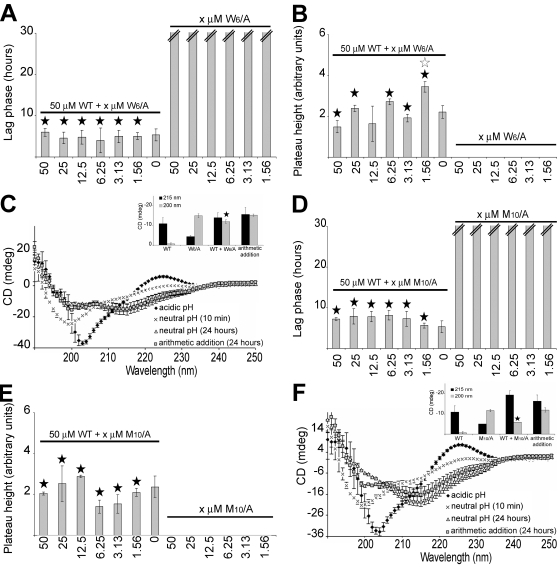
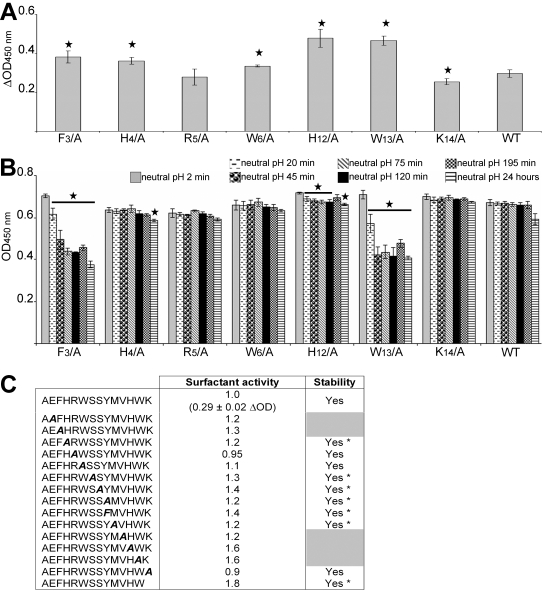
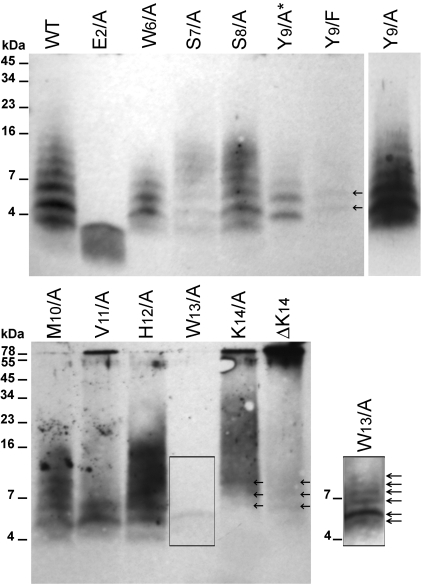
Polymerization into amyloid fibrils is a crucial step in the pathogenesis of neurodegenerative syndromes. Amyloid assembly is governed by properties of the sequence backbone and specific side-chain interactions, since fibrils from unrelated sequences possess similar structures and morphologies. Therefore, characterization of the structural determinants driving amyloid aggregation is of fundamental importance. We investigated the forces involved in the amyloid assembly of a model peptide derived from the oligomerization domain of acetylcholinesterase (AChE), AChE(586-599), through the effect of single point mutations on beta-sheet propensity, conformation, fibrilization, surfactant activity, oligomerization and fibril morphology. AChE(586-599) was chosen due to its fibrilization tractability and AChE involvement in Alzheimer's disease. The results revealed how specific regions and residues can control AChE(586-599) assembly. Hydrophobic and/or aromatic residues were crucial for maintaining a high beta-strand propensity, for the conformational transition to beta-sheet, and for the first stage of aggregation. We also demonstrated that positively charged side-chains might be involved in electrostatic interactions, which could control the transition to beta-sheet, the oligomerization and assembly stability. Further interactions were also found to participate in the assembly. We showed that some residues were important for AChE(586-599) surfactant activity and that amyloid assembly might preferentially occur at an air-water interface. Consistently with the experimental observations and assembly models for other amyloid systems, we propose a model for AChE(586-599) assembly in which a steric-zipper formed through specific interactions (hydrophobic, electrostatic, cation-pi, SH-aromatic, metal chelation and polar-polar) would maintain the beta-sheets together. We also propose that the stacking between the strands in the beta-sheets along the fiber axis could be stabilized through pi-pi interactions and metal chelation. The dissection of the specific molecular recognition driving AChE(586-599) amyloid assembly has provided further knowledge on such poorly understood and complicated process, which could be applied to protein folding and the targeting of amyloid diseases.
Conflict of interest statement
Figures
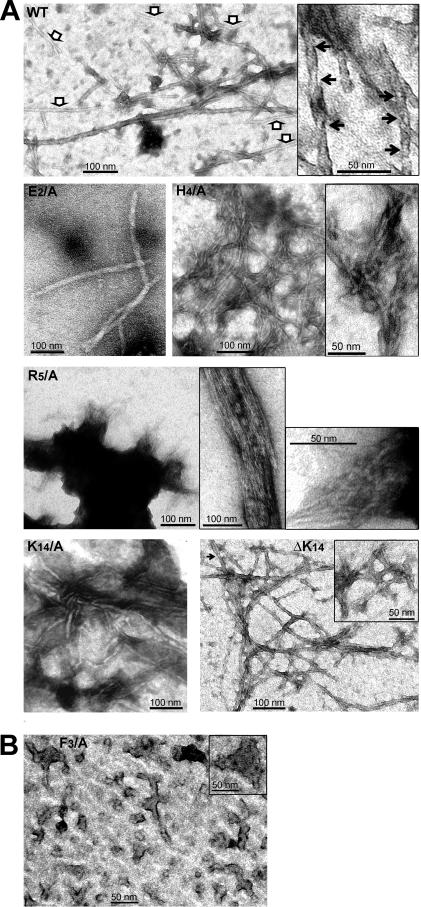
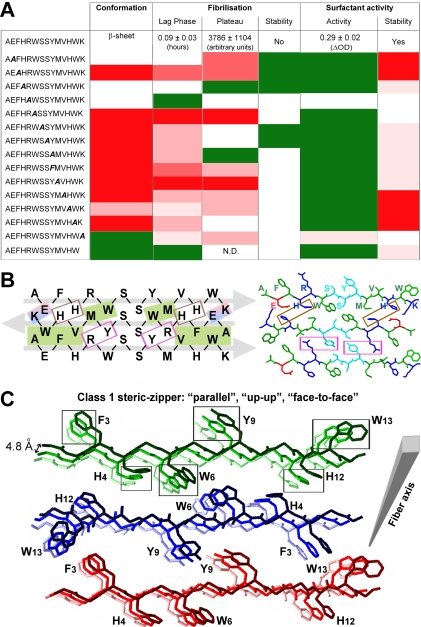
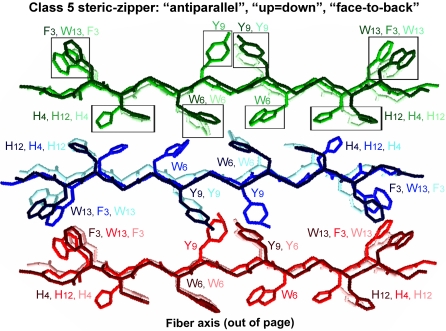
References
-
- Harper JD, Lansbury PT., Jr Models of amyloid seeding in Alzheimer's disease and scrapie: mechanistic truths and physiological consequences of the time-dependent solubility of amyloid proteins. Annu Rev Biochem. 1997;66:385–407. - PubMed
-
- Westermark P. Aspects on human amyloid forms and their fibril polypeptides. Febs J. 2005;272:5942–5949. - PubMed
-
- Klunk WE, Pettegrew JW, Abraham DJ. Quantitative evaluation of congo red binding to amyloid-like proteins with a beta-pleated sheet conformation. J Histochem Cytochem. 1989;37:1273–1281. - PubMed
-
- Rochet JC, Lansbury PT., Jr Amyloid fibrillogenesis: themes and variations. Curr Opin Struct Biol. 2000;10:60–68. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
