

Prenatal choline supplementation attenuates neuropathological response to status epilepticus in the adult rat hippocampus
- PMID: 18353663
- PMCID: PMC2413180
- DOI: 10.1016/j.nbd.2008.01.008
Prenatal choline supplementation attenuates neuropathological response to status epilepticus in the adult rat hippocampus
Abstract
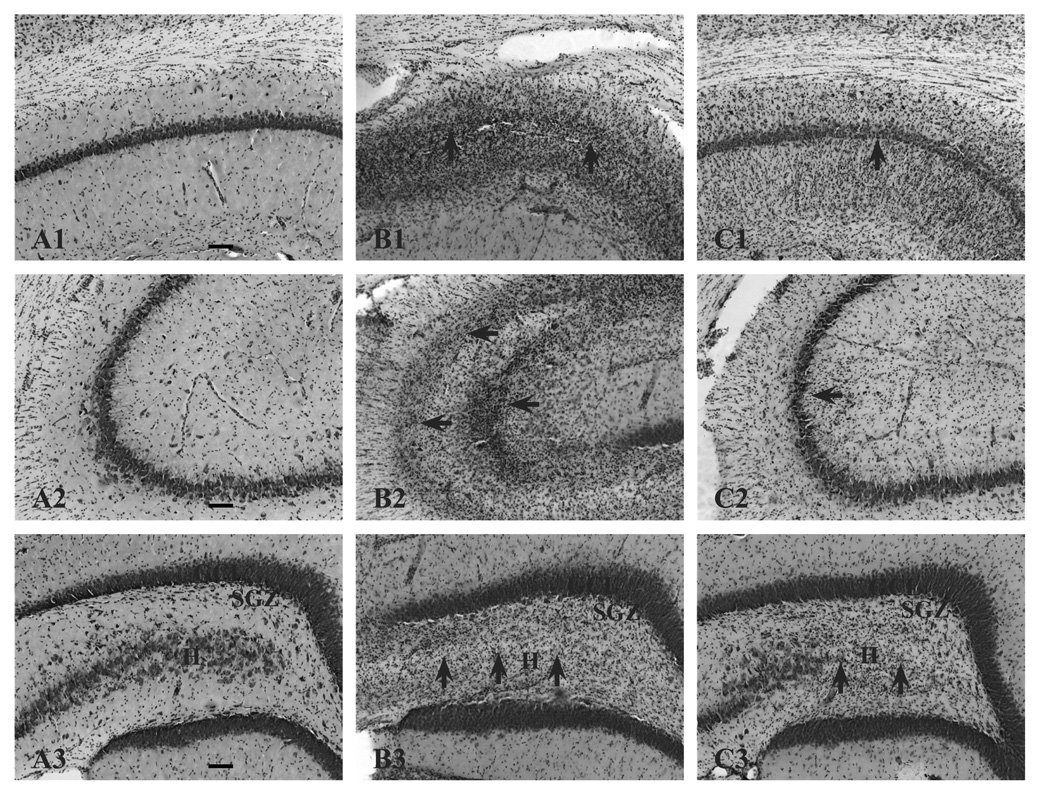
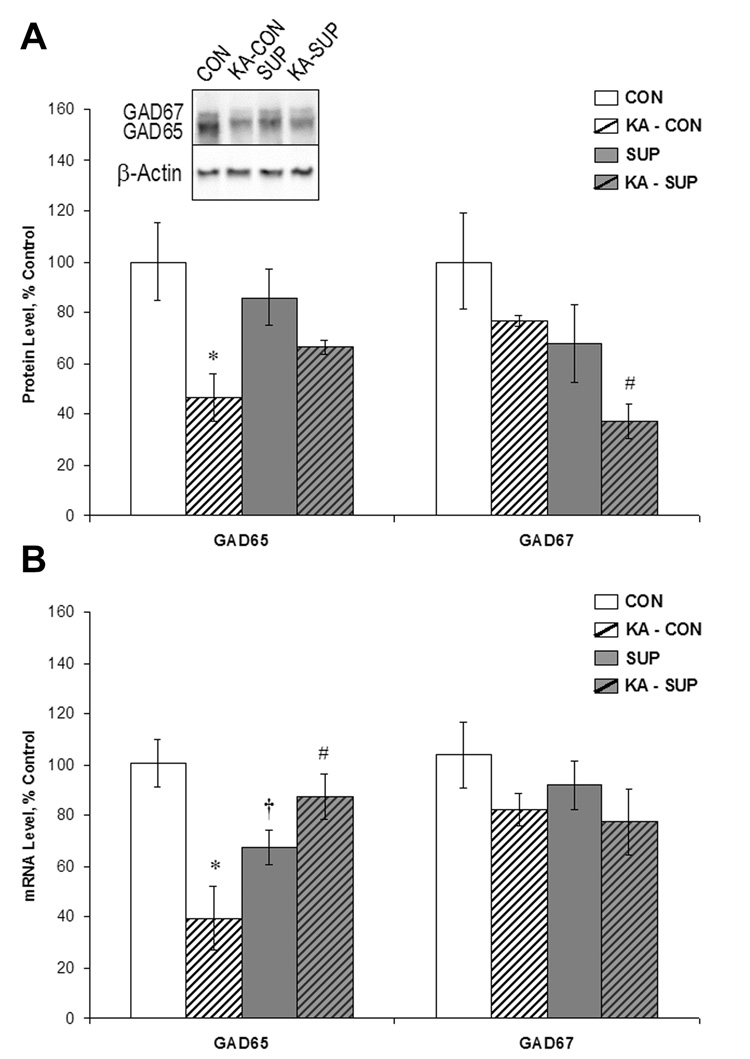
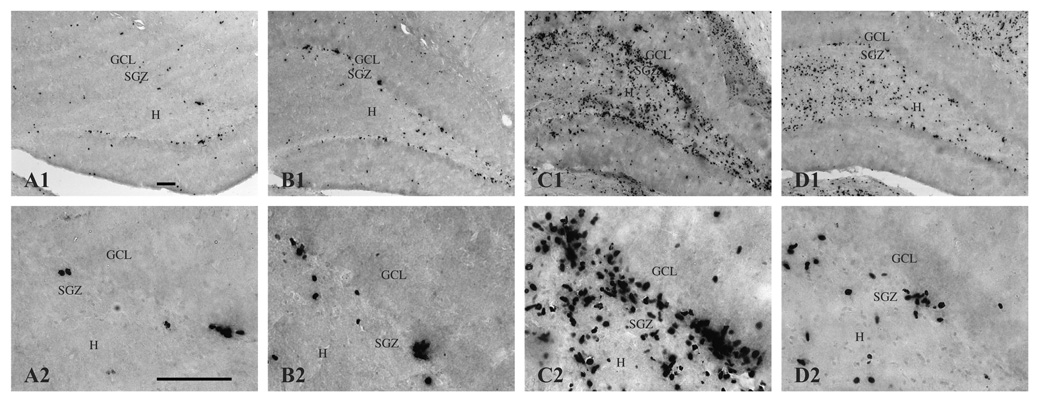
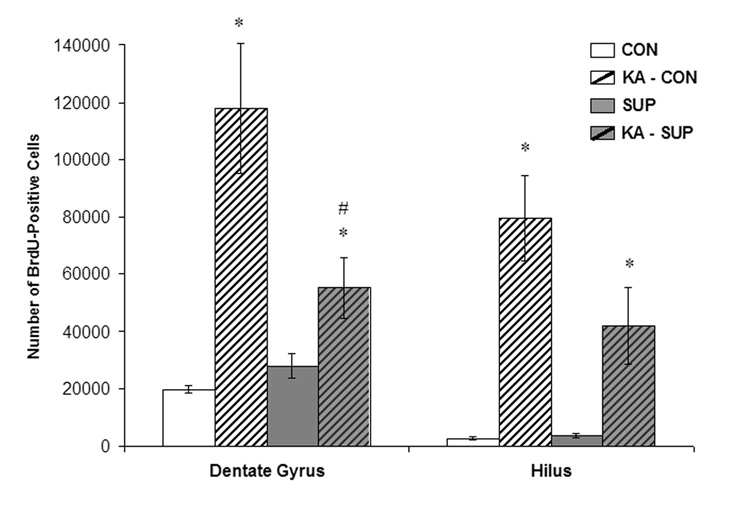
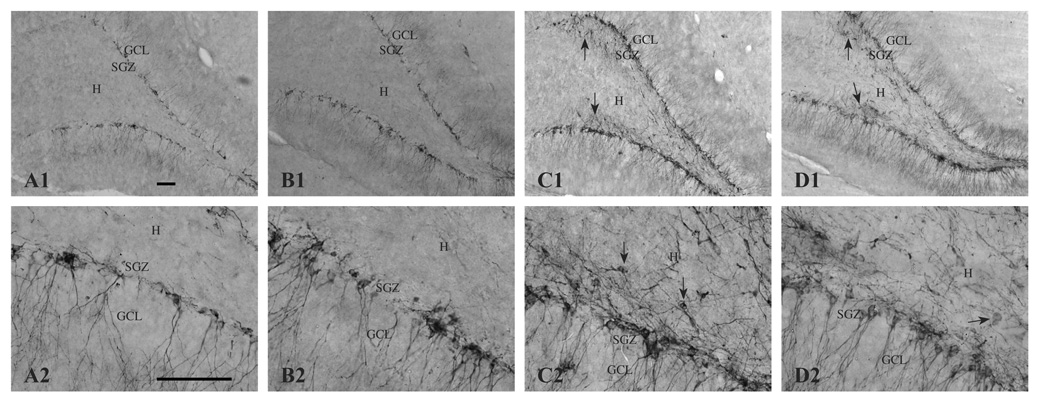
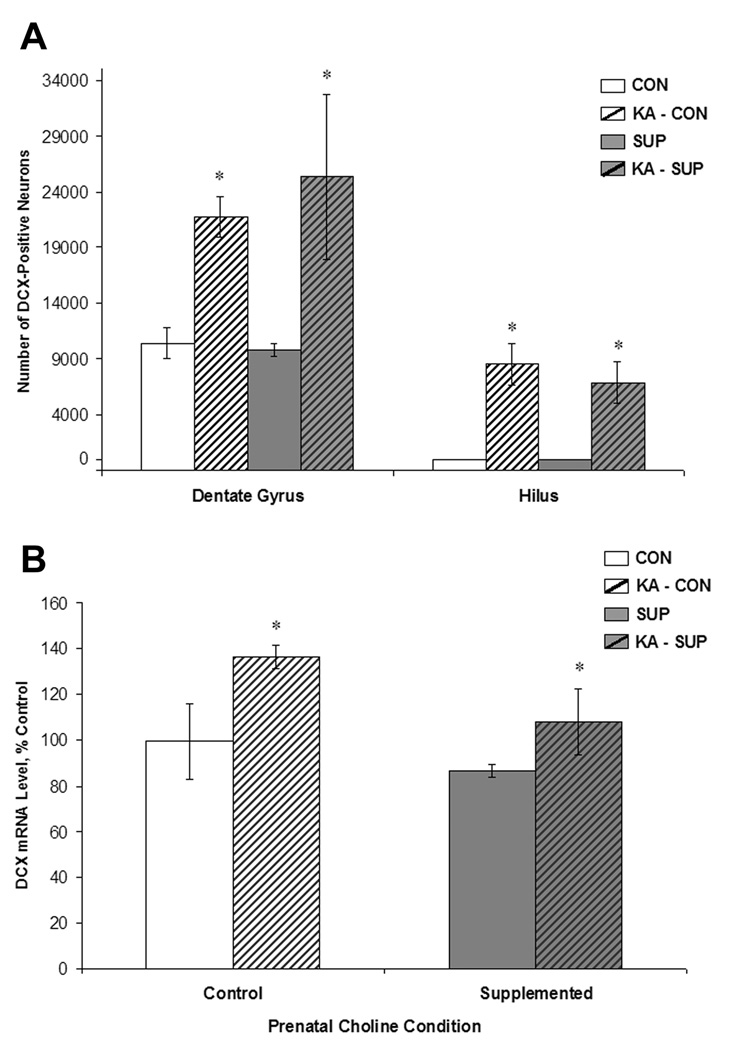
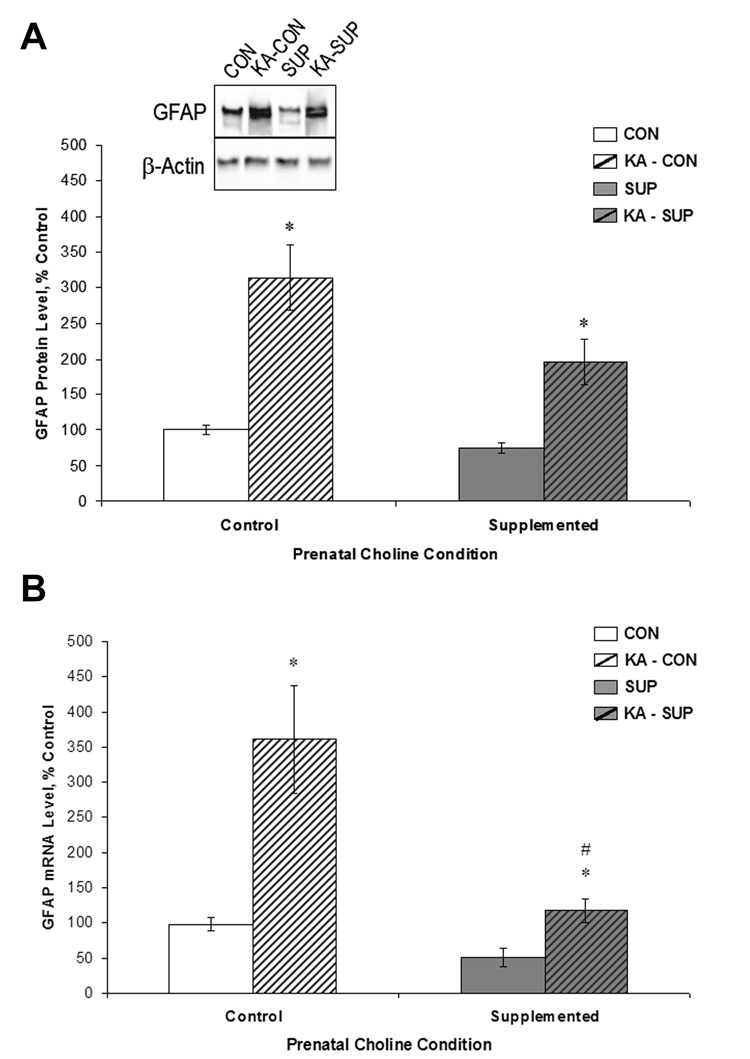
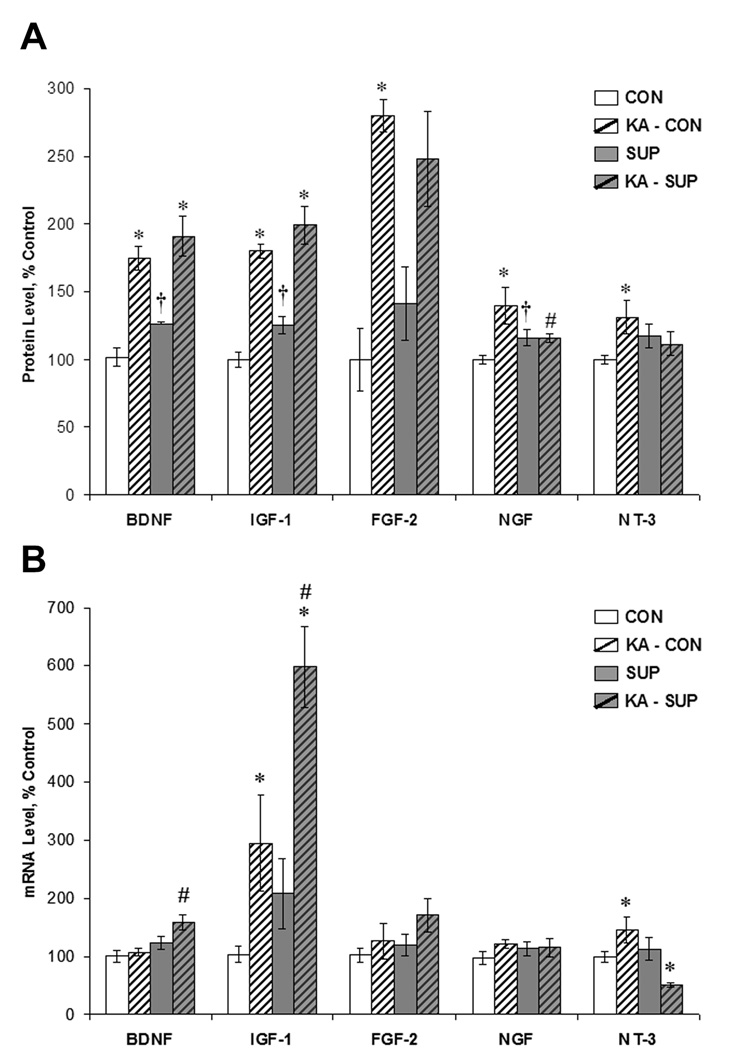
Prenatal choline supplementation (SUP) protects adult rats against spatial memory deficits observed after excitotoxin-induced status epilepticus (SE). To examine the mechanism underlying this neuroprotection, we determined the effects of SUP on a variety of hippocampal markers known to change in response to SE and thought to underlie ensuing cognitive deficits. Adult offspring from rat dams that received either a control or SUP diet on embryonic days 12-17 were administered saline or kainic acid (i.p.) to induce SE and were euthanized 16 days later. SUP markedly attenuated seizure-induced hippocampal neurodegeneration, dentate cell proliferation, and hippocampal GFAP mRNA expression levels, prevented the loss of hippocampal GAD65 protein and mRNA expression, and altered growth factor expression patterns. SUP also enhanced pre-seizure hippocampal levels of BDNF, NGF, and IGF-1, which may confer a neuroprotective hippocampal microenvironment that dampens the neuropathological response to and/or helps facilitate recovery from SE to protect cognitive function.
Figures
References
-
- Anderson MF, Aberg MA, Nilsson M, Eriksson PS. Insulin-like growth factor-I and neurogenesis in the adult mammalian brain. Brain Res Dev Brain Res. 2002;134:115–122. - PubMed
-
- Ben-Ari Y, Represa A. Brief seizure episodes induce long-term potentiation and mossy fiber sprouting in the hippocampus. Trends Neurosci. 1990;13:312–318. - PubMed
-
- Blusztajn JK. Choline, a vital amine. Science. 1998;281:794–795. - PubMed
-
- Blusztajn JK, Cermak JM, Holler T, Jackson DA. Imprinting of hippocampal metabolism of choline by its availability during gestation: implications for cholinergic neurotransmission. J Physiol Paris. 1998;92:199–203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
