

Design of a neuronal array
- PMID: 18354021
- PMCID: PMC2646167
- DOI: 10.1523/JNEUROSCI.5259-07.2008
Design of a neuronal array
Abstract
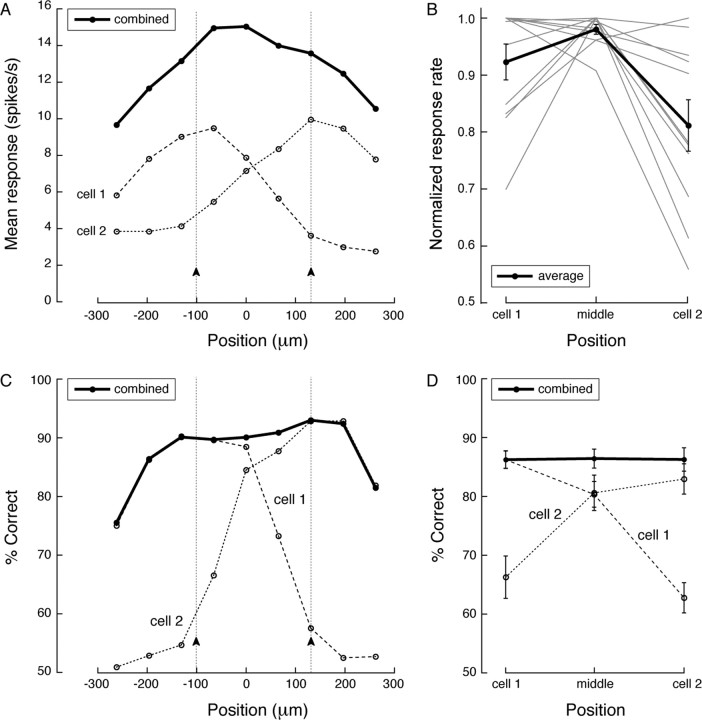
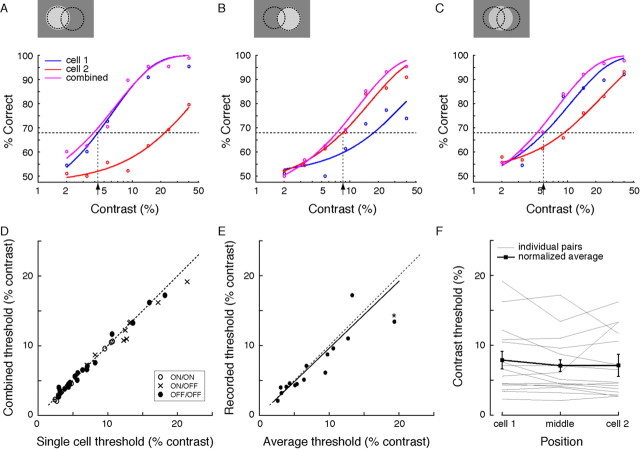
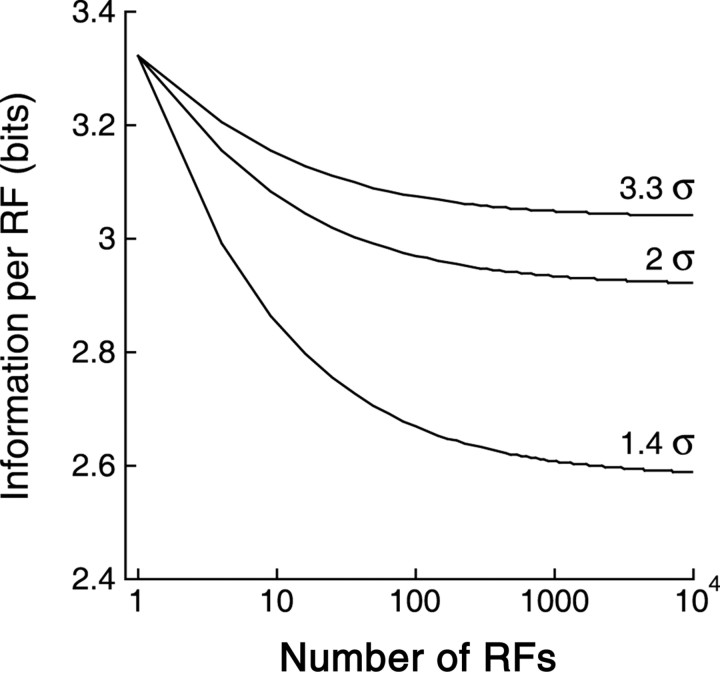
Retinal ganglion cells of a given type overlap their dendritic fields such that every point in space is covered by three to four cells. We investigated what function is served by such extensive overlap. Recording from pairs of ON or OFF brisk-transient ganglion cells at photopic intensities, we confirmed that this overlap causes the Gaussian receptive field centers to be spaced at approximately 2 SDs (sigma). This, together with response nonlinearities and variability, was just sufficient to provide an ideal observer with uniform contrast sensitivity across the retina for both threshold and suprathreshold stimuli. We hypothesized that overlap might maximize the information represented from natural images, thereby optimizing retinal performance for many tasks. Indeed, tested with natural images (which contain statistical correlations), a model ganglion cell array maximized information represented in its population responses with approximately 2sigma spacing, i.e., the overlap observed in the retina. Yet, tested with white noise (which lacks statistical correlations), an array maximized its information by minimizing overlap. In both cases, optimal overlap balanced greater signal-to-noise ratio (from larger receptive fields) against greater redundancy (because of larger receptive field overlap). Thus, dendritic overlap improves vision by taking optimal advantage of the statistical correlations of natural scenes.
Figures





References
-
- Adelson EH. Saturation and adaptation in the rod system. Vision Res. 1982;22:1299–1312. - PubMed
-
- Atick JJ, Redlich AN. Towards a theory of early visual processing. Neural Comput. 1990;2:308–320.
-
- Atick JJ, Li Z, Redlich AN. Understanding retinal color coding from first principles. Neural Comput. 1992;4:559–572.
-
- Attwell D, Laughlin SB. An energy budget for signaling in the grey matter of the brain. J Cereb Blood Flow Metab. 2001;21:1133–1145. - PubMed
-
- Balasubramanian V, Kimber D, Berry MJ., II Metabolically efficient information processing. Neural Comput. 2001;13:799–815. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous