

Dendritic cell-dependent inhibition of B cell proliferation requires CD22
- PMID: 18354178
- PMCID: PMC2728079
- DOI: 10.4049/jimmunol.180.7.4561
Dendritic cell-dependent inhibition of B cell proliferation requires CD22
Abstract
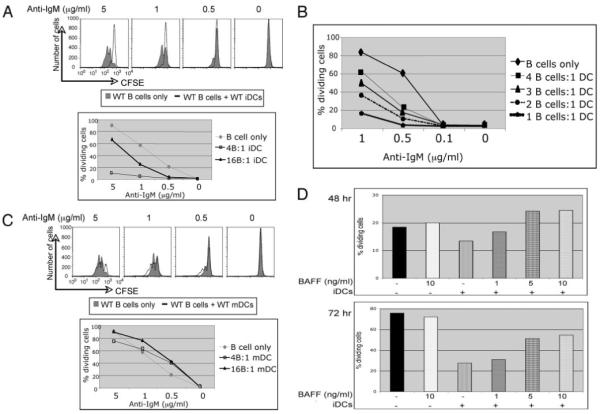
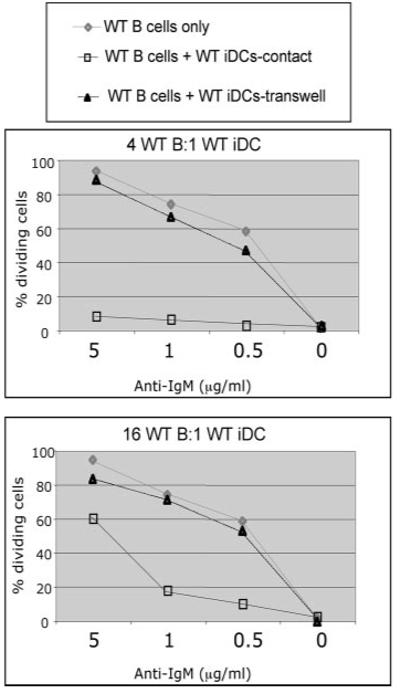
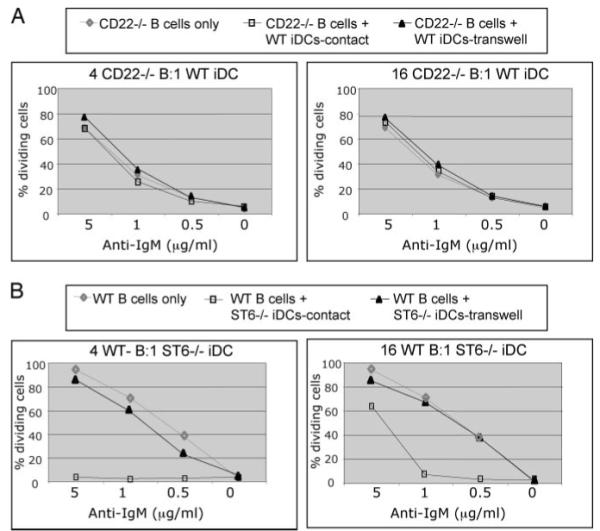
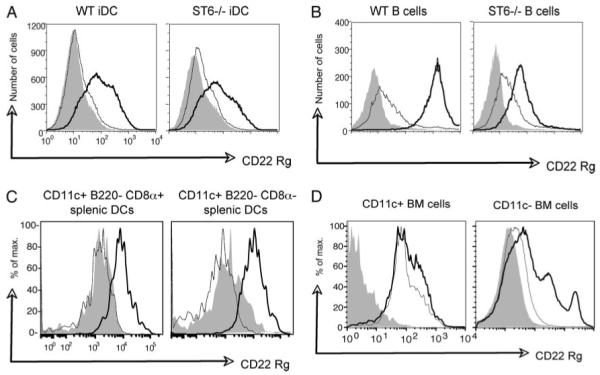
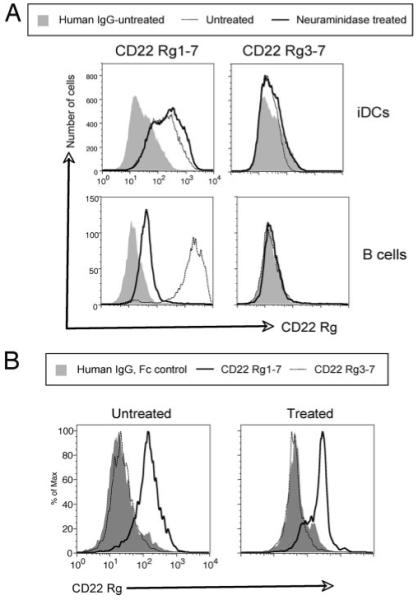
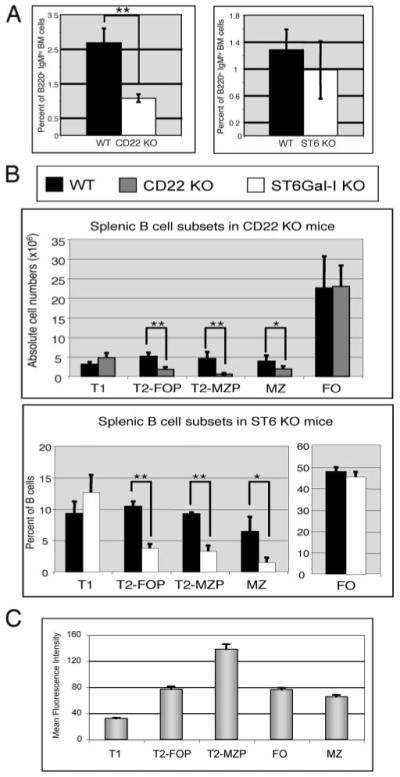
Recent studies have shown that dendritic cells (DCs) regulate B cell functions. In this study, we report that bone marrow (BM)-derived immature DCs, but not mature DCs, can inhibit BCR-induced proliferation of B cells in a contact-dependent manner. This inhibition is overcome by treatment with BAFF and is dependent on the BCR coreceptor CD22; however, it is not dependent on expression of the CD22 glycan ligand(s) produced by ST6Gal-I sialyltransferase. We found that a second CD22 ligand (CD22L) is expressed on CD11c(+) splenic and BM-derived DCs, which does not contain ST6Gal-I-generated sialic acids and which, unlike the B cell-associated CD22L, is resistant to neuraminidase treatment and sodium metaperiodate oxidation. Examination of splenic and BM B cell subsets in CD22 and ST6Gal-I knockout mice revealed that ST6Gal-I-generated B cell CD22L plays a role in splenic B cell development, whereas the maintenance of long-lived mature BM B cells depends only on CD22 and not on alpha2,6-sialic acids produced by ST6Gal-I. We propose that the two distinct CD22L have different functions. The alpha2,6-sialic acid-containing glycoprotein is important for splenic B cell subset development, whereas the DC-associated ST6Gal-I-independent CD22L may be required for the maintenance of long-lived mature B cells in the BM.
Figures
Similar articles
-
Ablation of CD22 in ligand-deficient mice restores B cell receptor signaling.Nat Immunol. 2006 Feb;7(2):199-206. doi: 10.1038/ni1283. Epub 2005 Dec 20. Nat Immunol. 2006. PMID: 16369536
-
Regulation of B cell development and B cell signalling by CD22 and its ligands alpha2,6-linked sialic acids.Int Immunol. 2006 Apr;18(4):603-11. doi: 10.1093/intimm/dxh402. Epub 2006 Feb 23. Int Immunol. 2006. PMID: 16497829
-
ST6Gal-I restrains CD22-dependent antigen receptor endocytosis and Shp-1 recruitment in normal and pathogenic immune signaling.Mol Cell Biol. 2006 Jul;26(13):4970-81. doi: 10.1128/MCB.00308-06. Mol Cell Biol. 2006. PMID: 16782884 Free PMC article.
-
CD22 and Siglec-G regulate inhibition of B-cell signaling by sialic acid ligand binding and control B-cell tolerance.Glycobiology. 2014 Sep;24(9):807-17. doi: 10.1093/glycob/cwu066. Epub 2014 Jul 6. Glycobiology. 2014. PMID: 25002414 Review.
-
B Cell Siglecs-News on Signaling and Its Interplay With Ligand Binding.Front Immunol. 2018 Dec 3;9:2820. doi: 10.3389/fimmu.2018.02820. eCollection 2018. Front Immunol. 2018. PMID: 30559744 Free PMC article. Review.
Cited by
-
Bone marrow dendritic cell-mediated regulation of TLR and B cell receptor signaling in B cells.J Immunol. 2012 Oct 1;189(7):3355-67. doi: 10.4049/jimmunol.1101352. Epub 2012 Aug 31. J Immunol. 2012. PMID: 22942427 Free PMC article.
-
Regulation of B-cell entry into the cell cycle.Immunol Rev. 2008 Aug;224:183-200. doi: 10.1111/j.1600-065X.2008.00652.x. Immunol Rev. 2008. PMID: 18759927 Free PMC article. Review.
-
LncRNA expression in idiopathic achalasia: New insight and preliminary exploration into pathogenesis.Open Med (Wars). 2022 Apr 12;17(1):732-740. doi: 10.1515/med-2022-0473. eCollection 2022. Open Med (Wars). 2022. PMID: 35509690 Free PMC article.
-
Macrophage regulation of B cell proliferation.Cell Immunol. 2017 Apr;314:54-62. doi: 10.1016/j.cellimm.2017.02.002. Epub 2017 Feb 21. Cell Immunol. 2017. PMID: 28238361 Free PMC article.
-
Coordinated Regulation of Extrafollicular B Cell Responses by IL-12 and IFNγ.Immunol Rev. 2025 May;331(1):e70027. doi: 10.1111/imr.70027. Immunol Rev. 2025. PMID: 40211749 Free PMC article. Review.
References
-
- Clark EA, Ledbetter JA. How B and T cells talk to each other. Nature. 1994;367:425–428. - PubMed
-
- Mellman I, Steinman RM. Dendritic cells: specialized and regulated antigen processing machines. Cell. 2001;106:255–258. - PubMed
-
- Kapsenberg ML. Dendritic-cell control of pathogen-driven T-cell polarization. Nat. Rev. Immunol. 2003;3:984–993. - PubMed
-
- Dubois B, Massacrier C, Vanbervliet B, Fayette J, Briere F, Banchereau J, Caux C. Critical role of IL-12 in dendritic cell-induced differentiation of naive B lymphocytes. J. Immunol. 1998;161:2223–2231. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
