

The initiation factor eIF3-f is a major target for atrogin1/MAFbx function in skeletal muscle atrophy
- PMID: 18354498
- PMCID: PMC2367397
- DOI: 10.1038/emboj.2008.52
The initiation factor eIF3-f is a major target for atrogin1/MAFbx function in skeletal muscle atrophy
Abstract
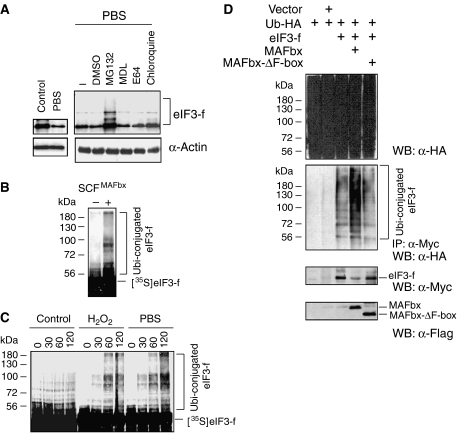
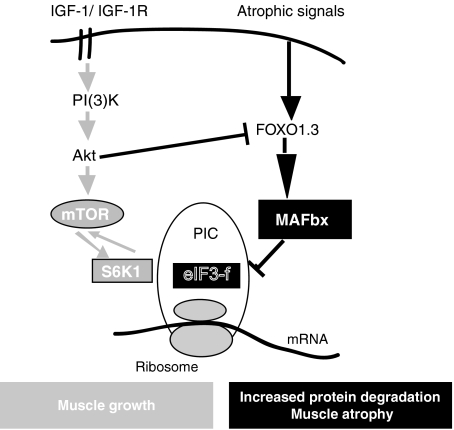
In response to cancer, AIDS, sepsis and other systemic diseases inducing muscle atrophy, the E3 ubiquitin ligase Atrogin1/MAFbx (MAFbx) is dramatically upregulated and this response is necessary for rapid atrophy. However, the precise function of MAFbx in muscle wasting has been questioned. Here, we present evidence that during muscle atrophy MAFbx targets the eukaryotic initiation factor 3 subunit 5 (eIF3-f) for ubiquitination and degradation by the proteasome. Ectopic expression of MAFbx in myotubes induces atrophy and degradation of eIF3-f. Conversely, blockade of MAFbx expression by small hairpin RNA interference prevents eIF3-f degradation in myotubes undergoing atrophy. Furthermore, genetic activation of eIF3-f is sufficient to cause hypertrophy and to block atrophy in myotubes, whereas genetic blockade of eIF3-f expression induces atrophy in myotubes. Finally, eIF3-f induces increasing expression of muscle structural proteins and hypertrophy in both myotubes and mouse skeletal muscle. We conclude that eIF3-f is a key target that accounts for MAFbx function during muscle atrophy and has a major role in skeletal muscle hypertrophy. Thus, eIF3-f seems to be an attractive therapeutic target.
Figures






Similar articles
-
MAFbx/Atrogin-1 controls the activity of the initiation factor eIF3-f in skeletal muscle atrophy by targeting multiple C-terminal lysines.J Biol Chem. 2009 Feb 13;284(7):4413-21. doi: 10.1074/jbc.M807641200. Epub 2008 Dec 10. J Biol Chem. 2009. PMID: 19073596
-
eIF3-f function in skeletal muscles: to stand at the crossroads of atrophy and hypertrophy.Cell Cycle. 2008 Jun 15;7(12):1698-701. doi: 10.4161/cc.7.12.6090. Epub 2008 Jun 11. Cell Cycle. 2008. PMID: 18583931
-
Suppression of atrogin-1 and MuRF1 prevents dexamethasone-induced atrophy of cultured myotubes.Metabolism. 2013 Oct;62(10):1495-502. doi: 10.1016/j.metabol.2013.05.018. Epub 2013 Jul 15. Metabolism. 2013. PMID: 23866982
-
Skeletal muscle atrophy and the E3 ubiquitin ligases MuRF1 and MAFbx/atrogin-1.Am J Physiol Endocrinol Metab. 2014 Sep 15;307(6):E469-84. doi: 10.1152/ajpendo.00204.2014. Epub 2014 Aug 5. Am J Physiol Endocrinol Metab. 2014. PMID: 25096180 Free PMC article. Review.
-
eIF3f: a central regulator of the antagonism atrophy/hypertrophy in skeletal muscle.Int J Biochem Cell Biol. 2013 Oct;45(10):2158-62. doi: 10.1016/j.biocel.2013.06.001. Epub 2013 Jun 13. Int J Biochem Cell Biol. 2013. PMID: 23769948 Free PMC article. Review.
Cited by
-
The ubiquitin ligase Nedd4-1 participates in denervation-induced skeletal muscle atrophy in mice.PLoS One. 2012;7(10):e46427. doi: 10.1371/journal.pone.0046427. Epub 2012 Oct 26. PLoS One. 2012. PMID: 23110050 Free PMC article.
-
Various jobs of proteolytic enzymes in skeletal muscle during unloading: facts and speculations.J Biomed Biotechnol. 2012;2012:493618. doi: 10.1155/2012/493618. Epub 2012 Feb 8. J Biomed Biotechnol. 2012. PMID: 22496611 Free PMC article. Review.
-
MAFbx/Atrogin-1 is required for atrophic remodeling of the unloaded heart.J Mol Cell Cardiol. 2014 Jul;72:168-76. doi: 10.1016/j.yjmcc.2014.03.006. Epub 2014 Mar 18. J Mol Cell Cardiol. 2014. PMID: 24650875 Free PMC article.
-
A potential role for Akt/FOXO signalling in both protein loss and the impairment of muscle carbohydrate oxidation during sepsis in rodent skeletal muscle.J Physiol. 2008 Nov 15;586(22):5589-600. doi: 10.1113/jphysiol.2008.160150. Epub 2008 Sep 25. J Physiol. 2008. PMID: 18818241 Free PMC article.
-
The effect of type 2 diabetes mellitus and obesity on muscle progenitor cell function.Stem Cell Res Ther. 2019 Mar 21;10(1):103. doi: 10.1186/s13287-019-1186-0. Stem Cell Res Ther. 2019. PMID: 30898146 Free PMC article. Review.
References
-
- Asano K, Phan L, Anderson J, Hinnebusch AG (1998) Complex formation by all five homologues of mammalian translation initiation factor 3 subunits from yeast Saccharomyces cerevisiae. J Biol Chem 27: 18573–18585 - PubMed
-
- Baar K, Esser K (1999) Phosphorylation of p70(S6k) correlates with increased skeletal muscle mass following resistance exercise. Am J Physiol 276: C120–C127 - PubMed
-
- Bodine SC, Latres E, Baumhueter S, Lai VK, Nunez L, Clarke BA, Poueymirou WT, Panaro FJ, Na E, Dharmarajan K, Pan ZQ, Valenzuela DM, DeChiara TM, Stitt TN, Yancopoulos GD, Glass DJ (2001a) Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 294: 1704–1708 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
