

Dissecting the role of putative CD81 binding regions of E2 in mediating HCV entry: putative CD81 binding region 1 is not involved in CD81 binding
- PMID: 18355410
- PMCID: PMC2277408
- DOI: 10.1186/1743-422X-5-46
Dissecting the role of putative CD81 binding regions of E2 in mediating HCV entry: putative CD81 binding region 1 is not involved in CD81 binding
Abstract
Background: Hepatitis C virus (HCV) encodes two transmembrane glycoproteins E1 and E2 which form a heterodimer. E1 is believed to mediate fusion while E2 has been shown to bind cellular receptors including CD81. In this study, alanine substitutions in E2 were generated within putative CD81 binding regions to define residues critical for viral entry. The effect of each mutation was tested by challenging susceptible cell lines with mutant HCV E1E2 pseudotyped viruses generated using a lentiviral system (HCVpp). In addition to assaying infectivity, producer cell expression and HCVpp incorporation of HCV E1 and E2 proteins, CD81 binding profiles, and E1E2 association of mutants were examined.
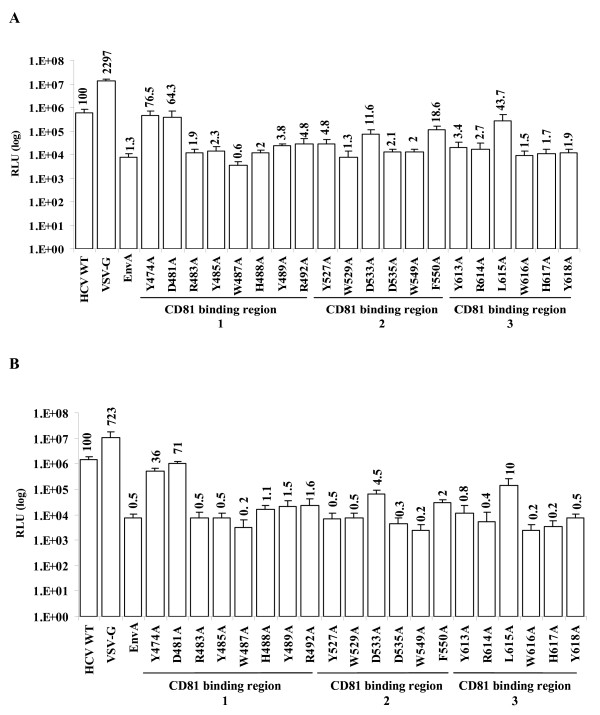
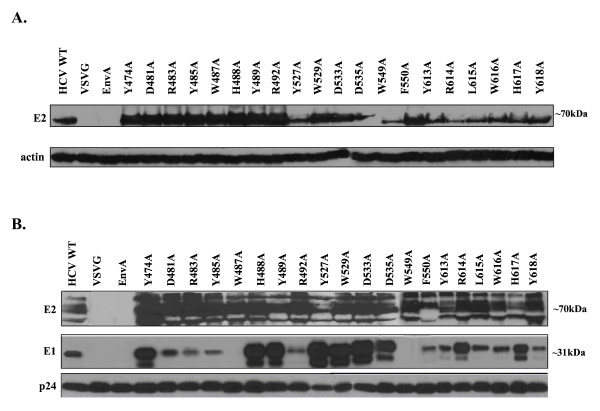
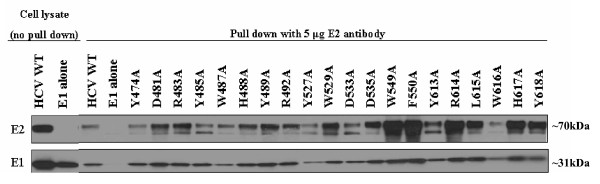
Results: Based on these characteristics, mutants either displayed wt characteristics (high infectivity [> or = 50% of wt HCVpp], CD81 binding, E1E2 expression, association, and incorporation into viral particles and proper conformation) or segregated into 4 distinct low infectivity (< or = 50% of wt HCVpp) mutant phenotypes: (I) CD81 binding deficient (despite wt E1E2 expression, incorporation and association and proper conformation); (II) CD81 binding competent, but lack of E1 detection on the viral particle, (despite adequate E1E2 expression in producer cell lysates and proper conformation); (III) CD81 binding competent, with adequate E1E2 expression, incorporation, association, and proper E2 conformation (i.e. no defect identified to explain the reduced infectivity observed); (IV) CD81 binding deficient due to disruption of E2 mutant protein conformation.
Conclusion: Although most alanine substitutions within the putative CD81 binding region 1 (amino acids 474-492) displayed greatly reduced HCVpp infectivity, they retained soluble CD81 binding, proper E2 conformation, E1E2 association and incorporation into HCVpp suggesting that region 1 of E2 does not mediate binding to CD81. In contrast, conformationally correct E2 mutants (Y527 and W529) within the second putative CD81 binding region (amino acids 522-551) disrupted binding of E2 to CD81-GST, suggesting that region 2 is critical to CD81 binding. Likewise, all conformationally intact mutants within the third putative CD81 binding region (amino acids 612-619), except L615A, were important for E2 binding to CD81-GST. This region is highly conserved across genotypes, underlining its importance in mediating viral entry.
Figures


References
-
- Lindenbach BD, Rice CM. Flaviviridae: The viruses and their replication. In: Knipe DM and Howley PM, editor. Field's Virology. 4th. Vol. 2. Philadelphia, Pippincott Williams & Wilkins; 2001. pp. 991–1041.
-
- Lavie M, Goffard A, Dubuisson J. Assembly of a functional HCV glycoprotein heterodimer. Curr Issues Mol Biol. 2007;9:71–86. - PubMed
-
- Meola A, Sbardellati A, Bruni Ercole B, Cerretani M, Pezzanera M, Ceccacci A, Vitelli A, Levy S, Nicosia A, Traboni C, McKeating J, Scarselli E. Binding of hepatitis C virus E2 glycoprotein to CD81 does not correlate with species permissiveness to infection. J Virol. 2000;74:5933–5938. doi: 10.1128/JVI.74.13.5933-5938.2000. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
