

A novel role for zebrafish zic2a during forebrain development
- PMID: 18358464
- PMCID: PMC2409112
- DOI: 10.1016/j.ydbio.2008.02.029
A novel role for zebrafish zic2a during forebrain development
Abstract
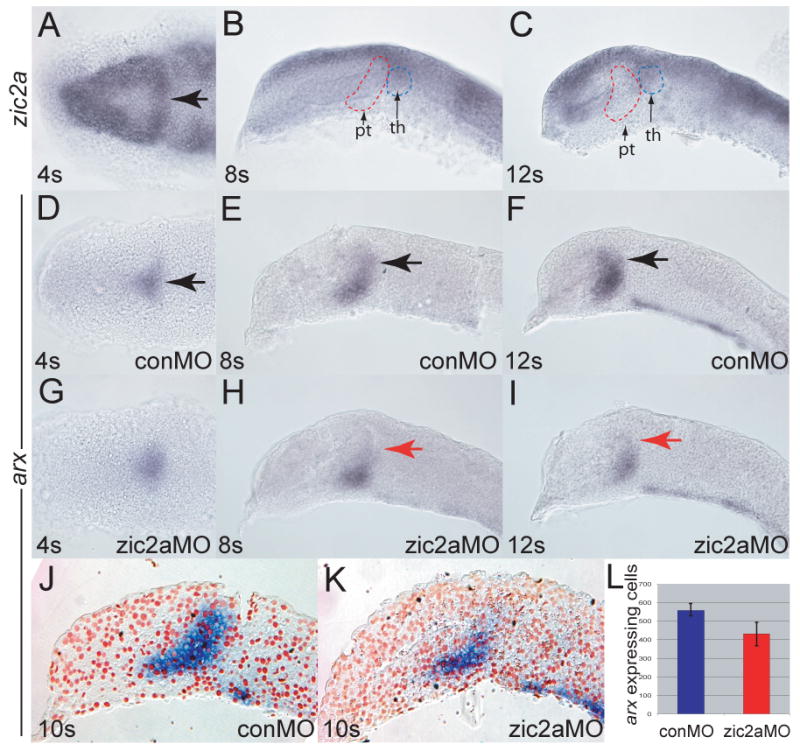
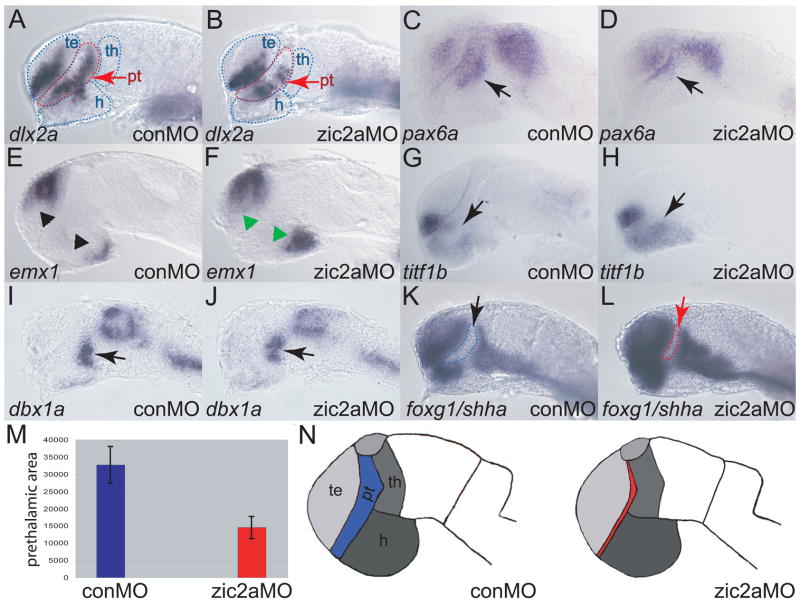
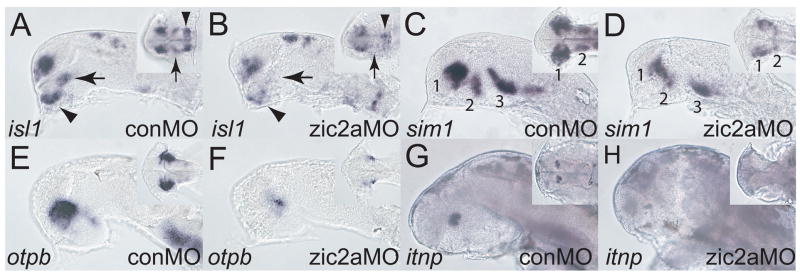
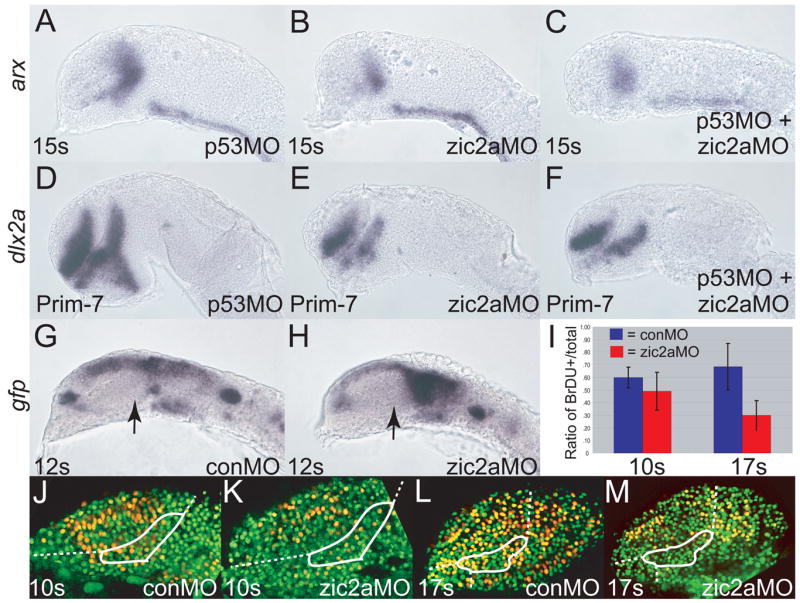
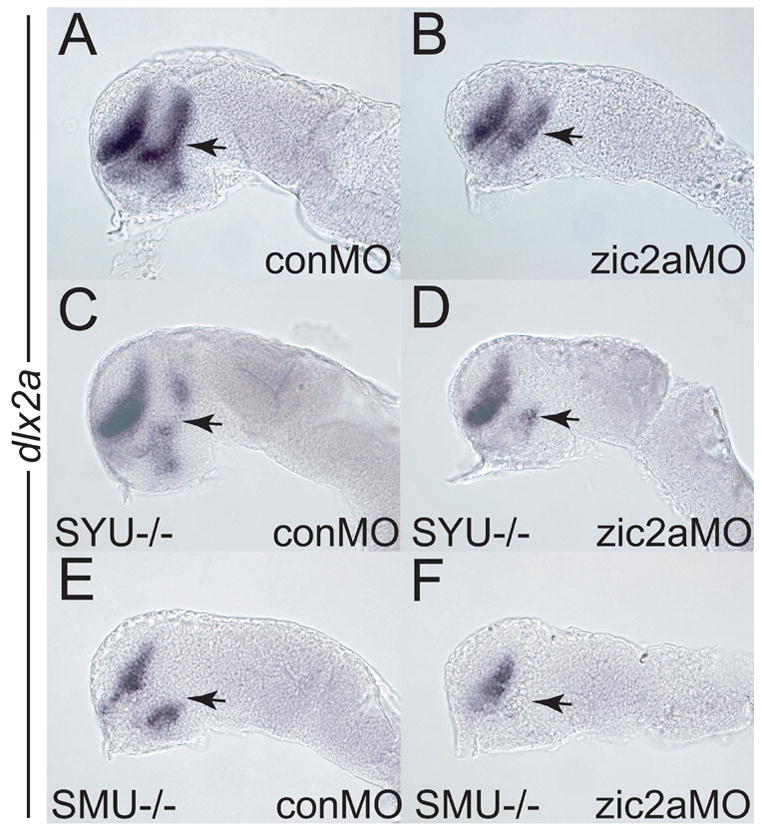
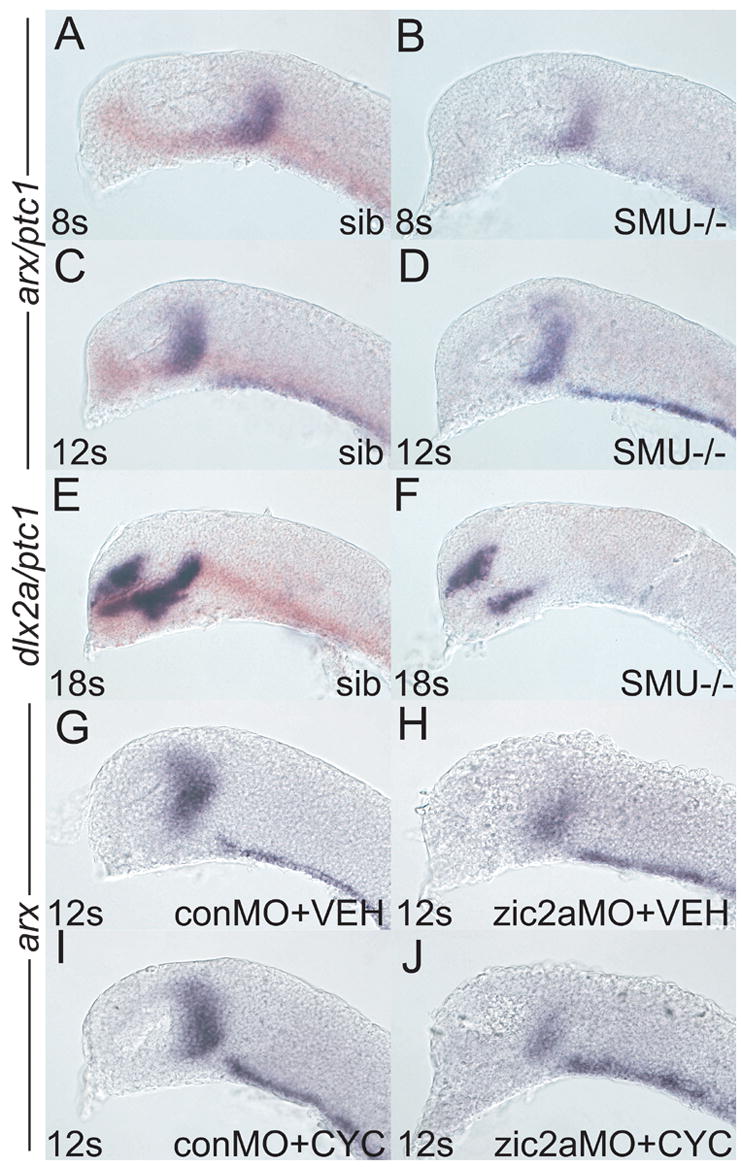
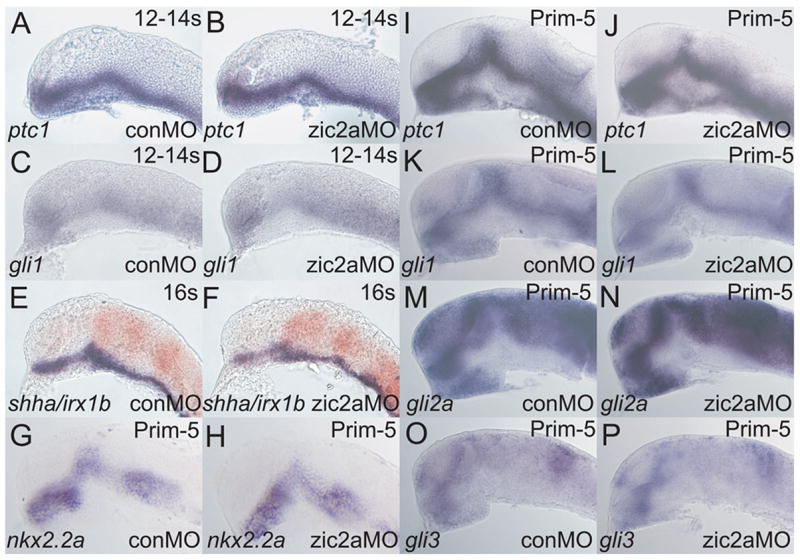
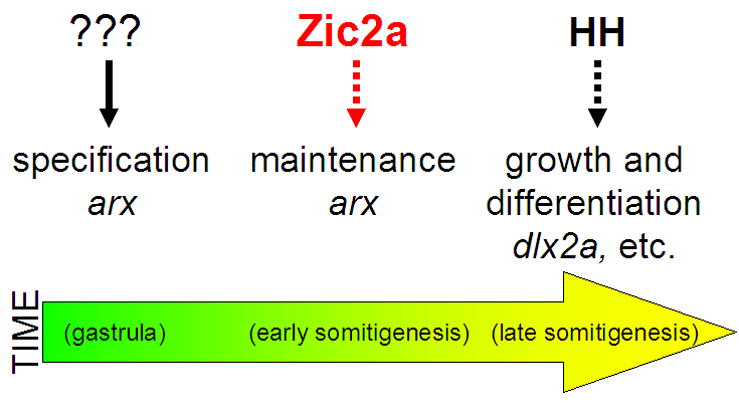
Patterns of transcription factor expression establish a blueprint for the vertebrate forebrain early in embryogenesis. In the future diencephalon, several genes with patterned expression have been identified, yet their specific functions and interactions between them are not well understood. We have uncovered a crucial role for one such gene, zic2a, during formation of the anterior diencephalon in zebrafish. We show that zic2a is required for transcription of the prethalamic markers arx and dlx2a. This function is required during early steps of prethalamic development, soon after its specification. zic genes are evolutionarily related to glis, transcription factors that mediate hedgehog signaling. Intriguingly, the hedgehog signaling pathway also acts to promote development of the prethalamus. We asked if zic2a interacts with hedgehog signaling in the context of forebrain development in zebrafish. Our data show that hedgehog signaling and zic2a function at different times, and therefore act in parallel pathways during forebrain development. Taken together, our results identify Zic2a as a novel regulator of prethalamic development, and show that it functions independently of hedgehog signaling.
Figures
References
-
- Amores A, Force A, Yan YL, Joly L, Amemiya C, Fritz A, Ho RK, Langeland J, Prince V, Wang YL, Westerfield M, Ekker M, Postlethwait JH. Zebrafish hox clusters and vertebrate genome evolution. Science. 1998;282:1711–4. - PubMed
-
- Aruga J. The role of Zic genes in neural development. Mol Cell Neurosci. 2004;26:205–21. - PubMed
-
- Aruga J, Mizugishi K, Koseki H, Imai K, Balling R, Noda T, Mikoshiba K. Zic1 regulates the patterning of vertebral arches in cooperation with Gli3. Mech Dev. 1999;89:141–50. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
