

Regulation of gene expression in a mixed-genus community: stabilized arginine biosynthesis in Streptococcus gordonii by coaggregation with Actinomyces naeslundii
- PMID: 18359813
- PMCID: PMC2395002
- DOI: 10.1128/JB.00088-08
Regulation of gene expression in a mixed-genus community: stabilized arginine biosynthesis in Streptococcus gordonii by coaggregation with Actinomyces naeslundii
Abstract
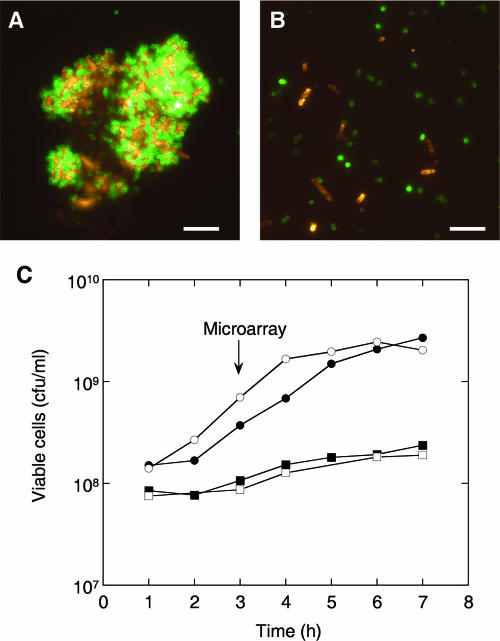
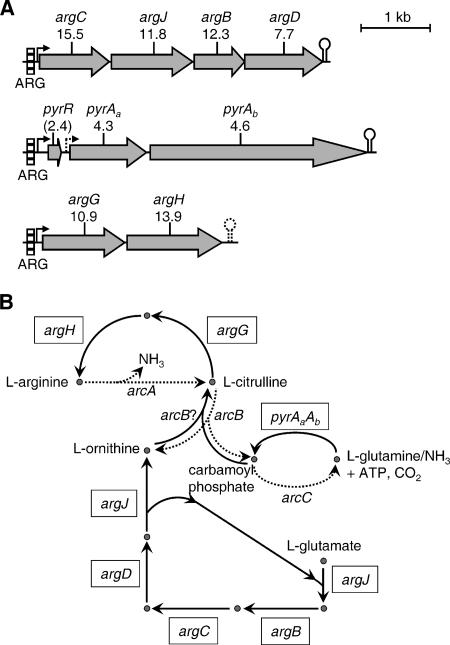
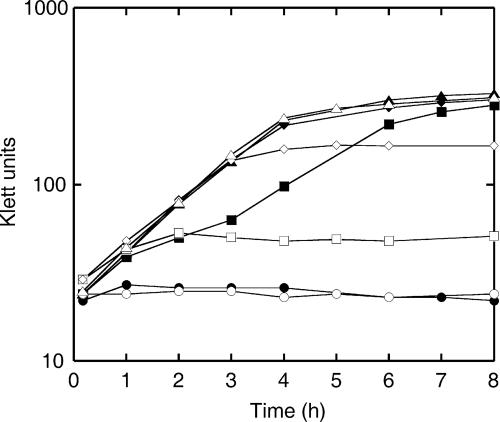
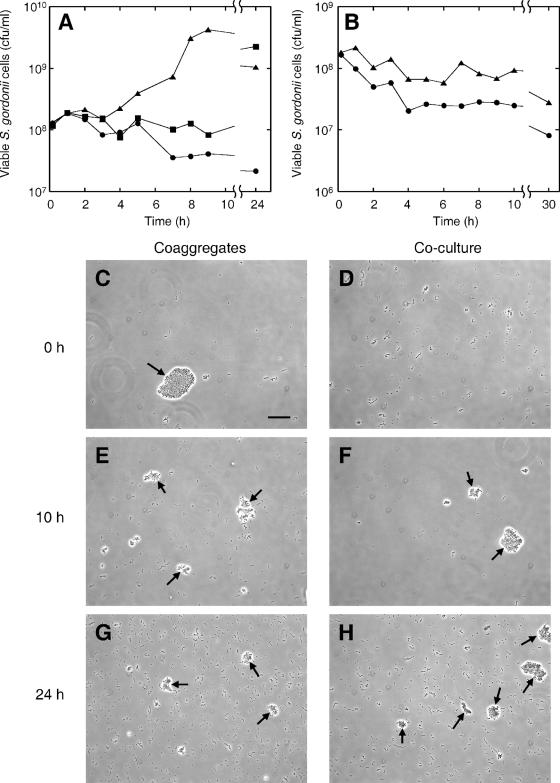
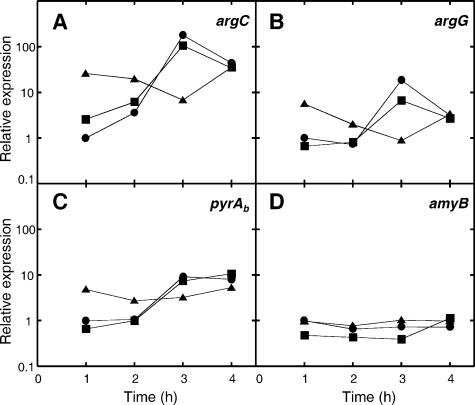
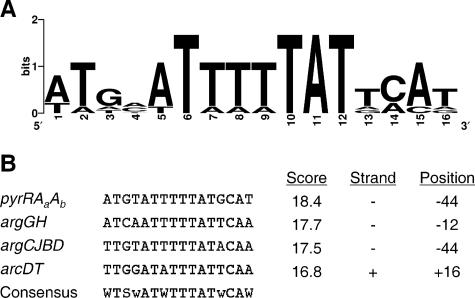
Interactions involving genetically distinct bacteria, for example, between oral streptococci and actinomyces, are central to dental plaque development. A DNA microarray identified Streptococcus gordonii genes regulated in response to coaggregation with Actinomyces naeslundii. The expression of 23 genes changed >3-fold in coaggregates, including that of 9 genes involved in arginine biosynthesis and transport. The capacity of S. gordonii to synthesize arginine was assessed using a chemically defined growth medium. In monoculture, streptococcal arginine biosynthesis was inefficient and streptococci could not grow aerobically at low arginine concentrations. In dual-species cultures containing coaggregates, however, S. gordonii grew to high cell density at low arginine concentrations. Equivalent cocultures without coaggregates showed no growth until coaggregation was evident (9 h). An argH mutant was unable to grow at low arginine concentrations with or without A. naeslundii, indicating that arginine biosynthesis was essential for coaggregation-induced streptococcal growth. Using quantitative reverse transcriptase PCR, the expression of argC, argG, and pyrA(b) was strongly (10- to 100-fold) up-regulated in S. gordonii monocultures after 3 h of growth when exogenous arginine was depleted. Cocultures without induced coaggregation showed similar regulation. However, within 1 h after coaggregation with A. naeslundii, the expression of argC, argG, and pyrA(b) in S. gordonii was partially up-regulated although arginine was plentiful, and mRNA levels did not increase further when arginine was diminished. Thus, A. naeslundii stabilizes S. gordonii expression of arginine biosynthesis genes in coaggregates but not cocultures and enables aerobic growth when exogenous arginine is limited.
Figures
References
-
- Brand, H. S., G. G. Jörning, R. A. Chamuleau, and L. Abraham-Inpijn. 1997. Effect of a protein-rich meal on urinary and salivary free amino acid concentrations in human subjects. Clin. Chim. Acta 26437-47. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
