

Role of flavin-containing monooxygenase in oxidative metabolism of voriconazole by human liver microsomes
- PMID: 18362161
- PMCID: PMC2737669
- DOI: 10.1124/dmd.107.019646
Role of flavin-containing monooxygenase in oxidative metabolism of voriconazole by human liver microsomes
Abstract
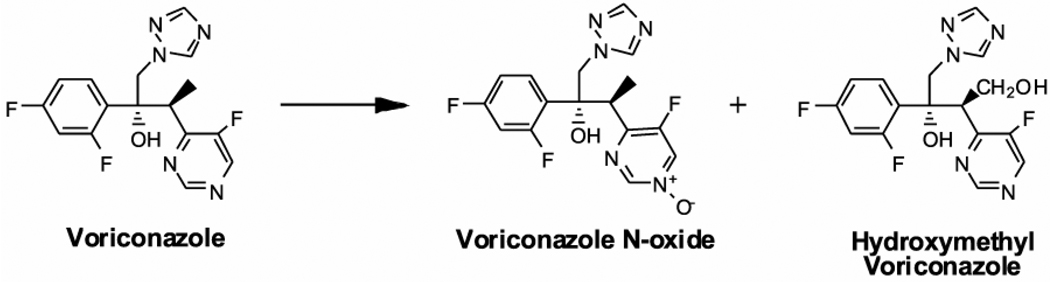
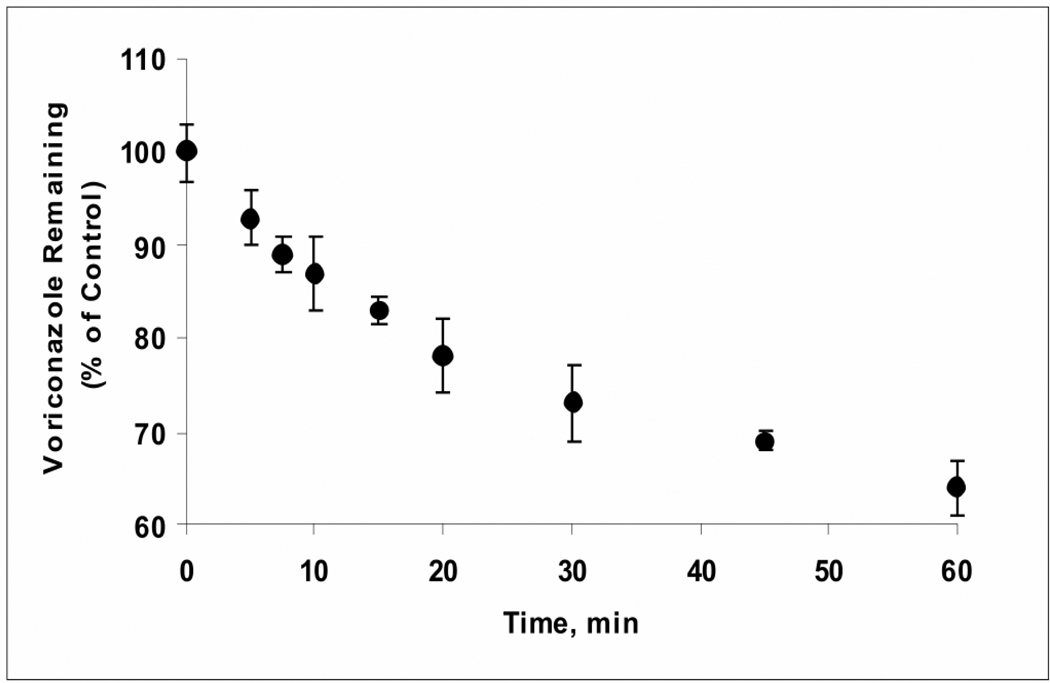
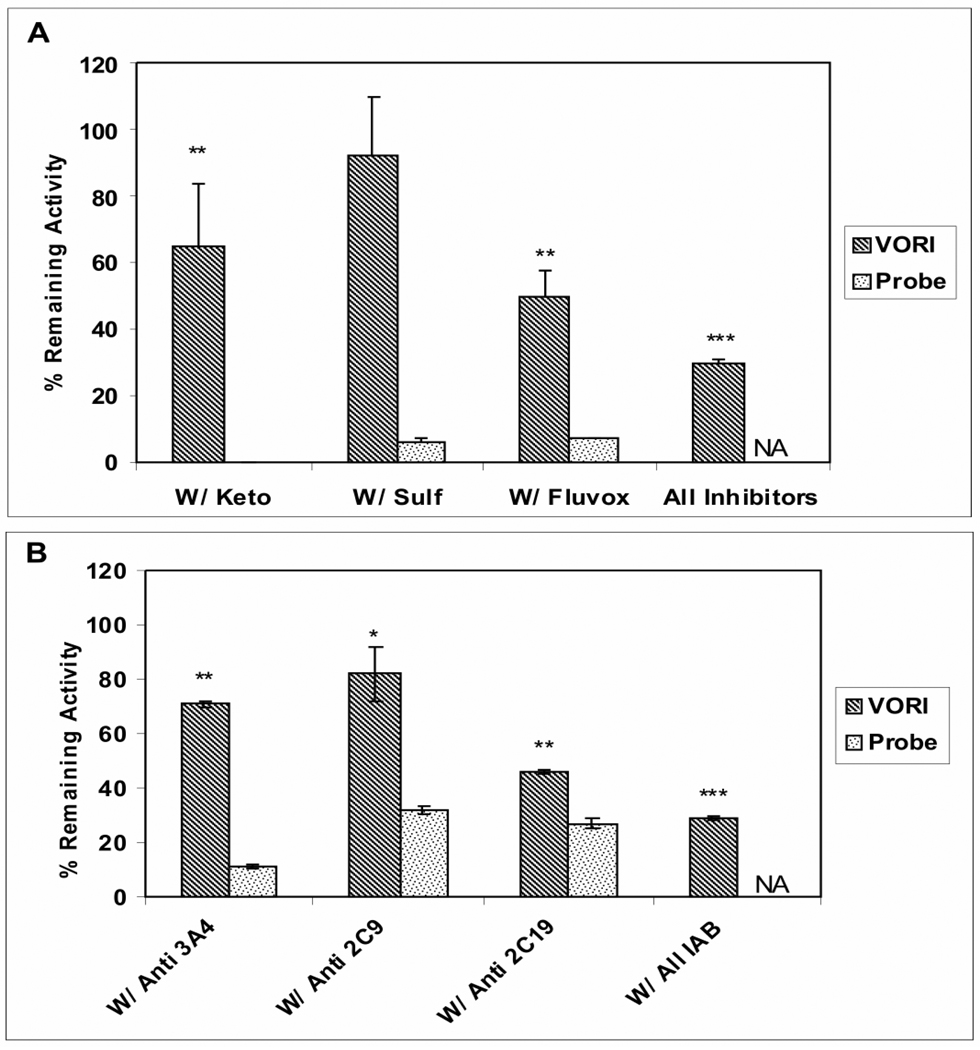
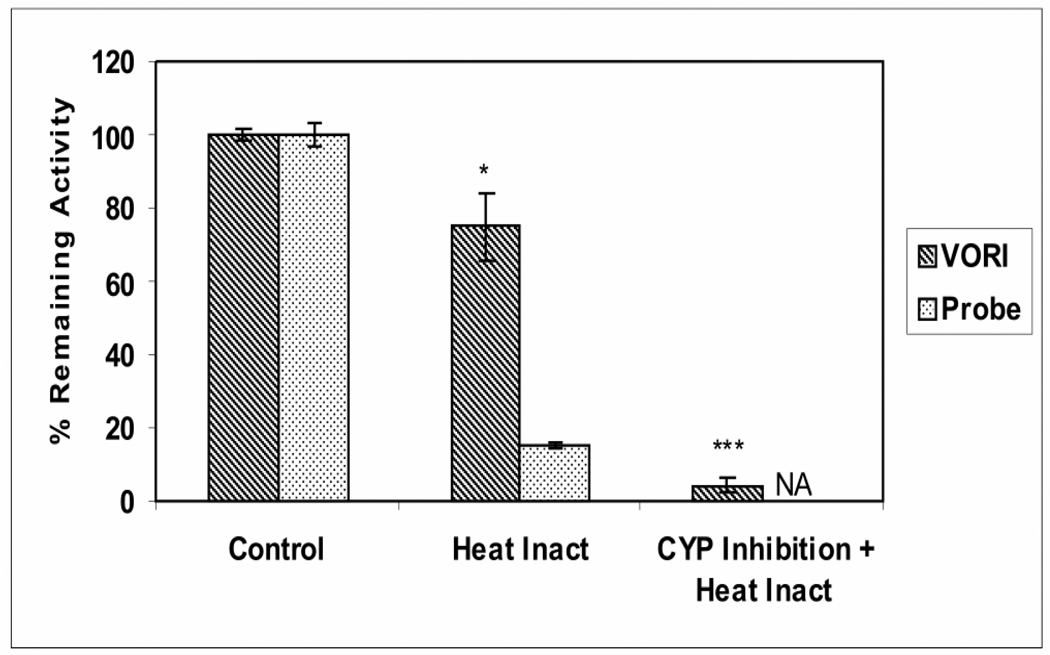
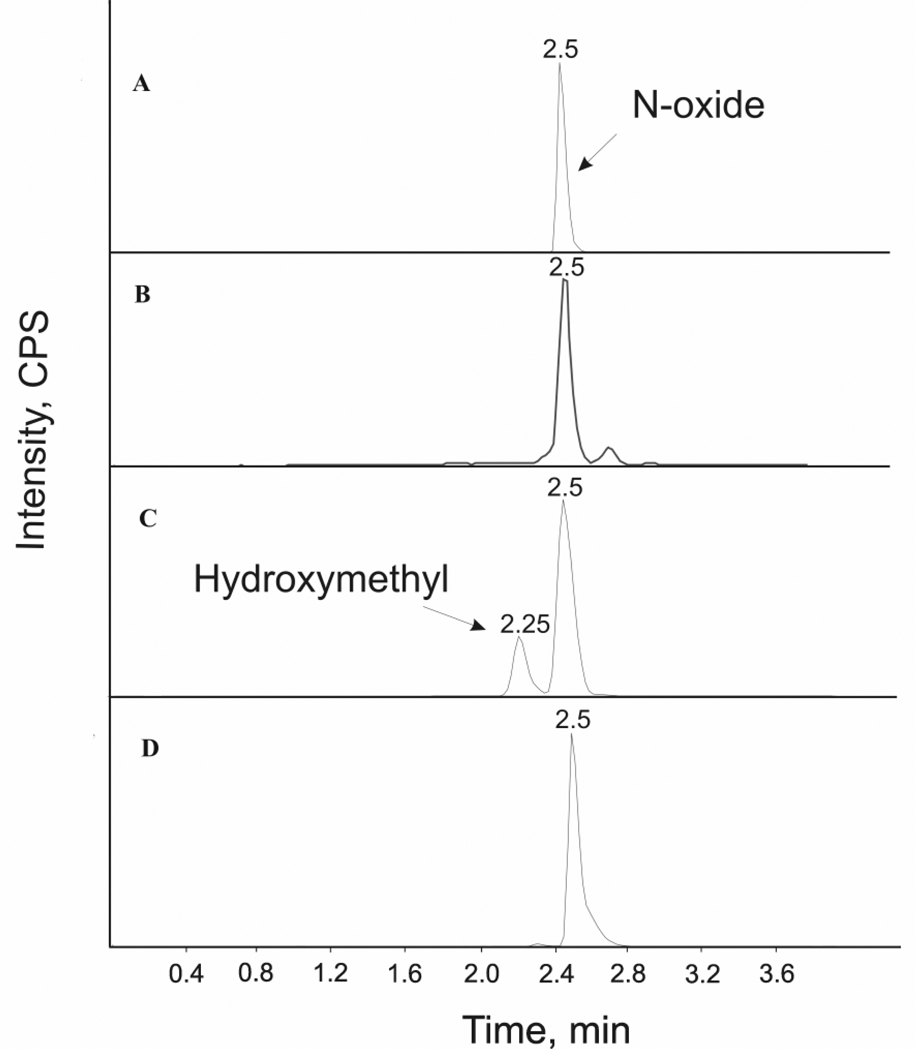
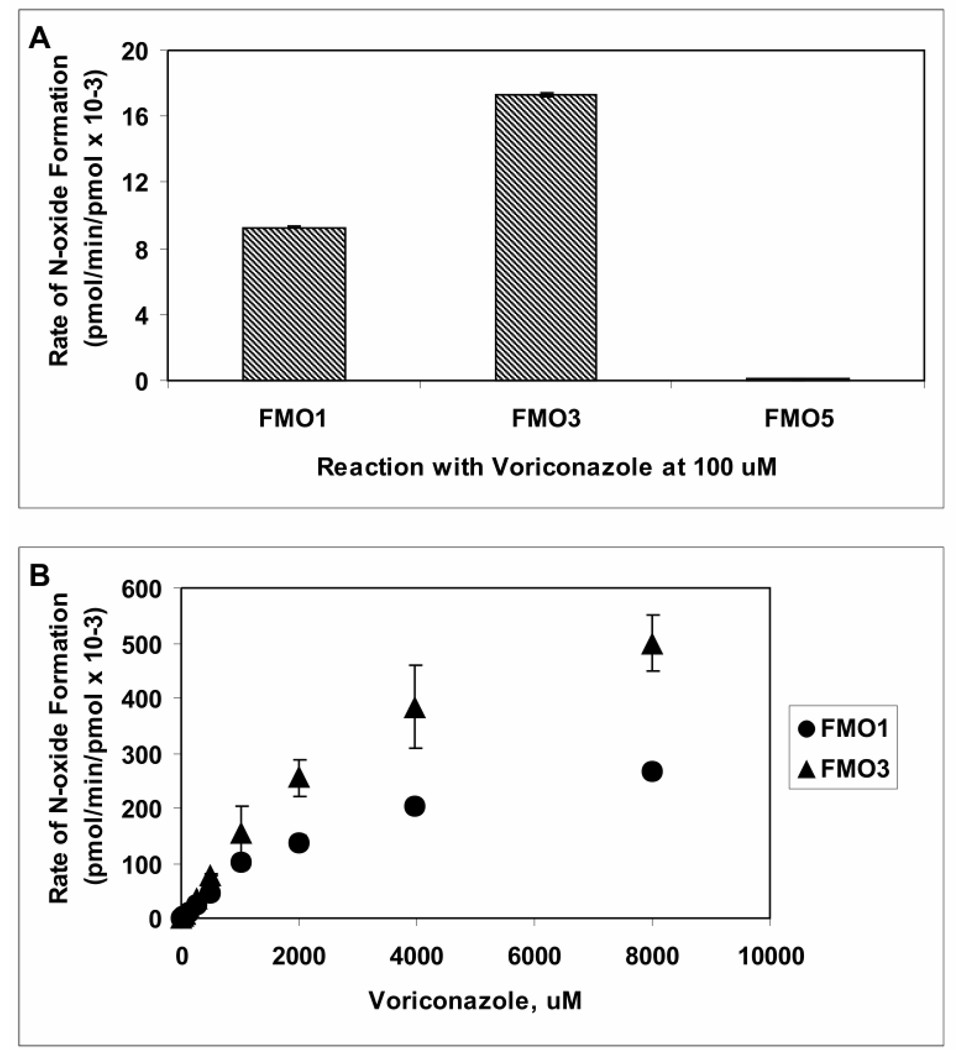
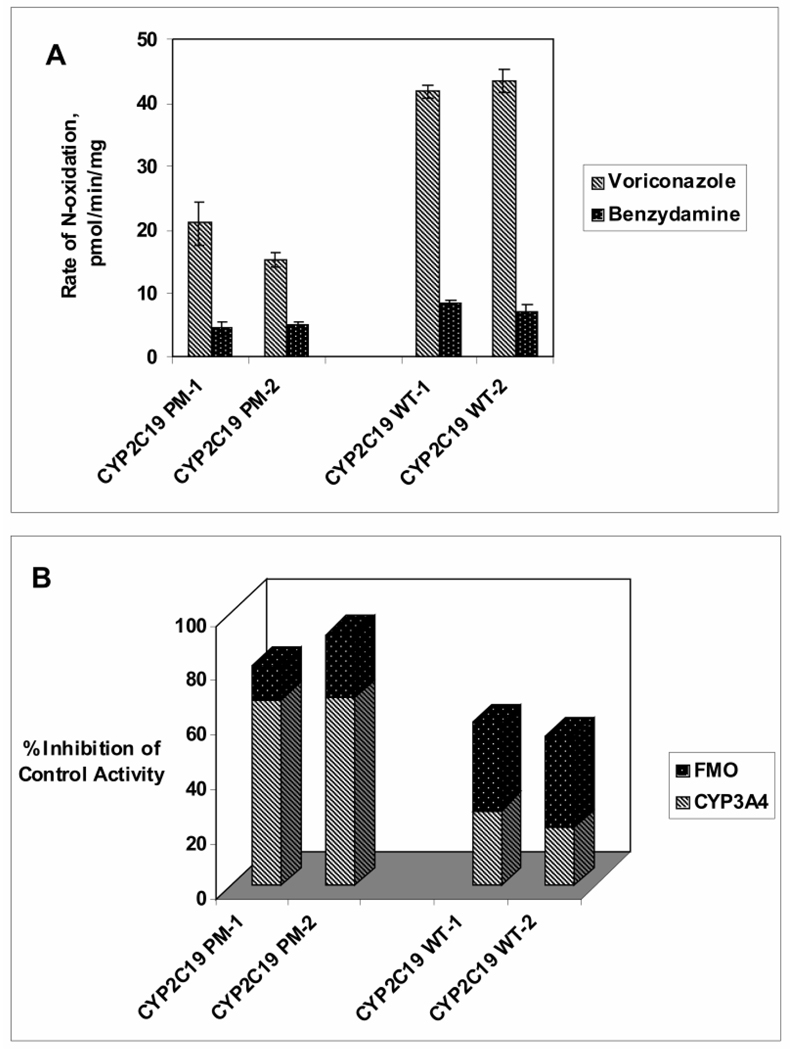
Voriconazole is a potent second-generation triazole antifungal agent with broad-spectrum activity against clinically important fungi. It is cleared predominantly via metabolism in all species tested including humans. N-Oxidation of the fluoropyrimidine ring, its hydroxylation, and hydroxylation of the adjacent methyl group are the known pathways of voriconazole oxidative metabolism, with the N-oxide being the major circulating metabolite in human. In vitro studies have shown that CYP2C19, CYP3A4, and to a lesser extent CYP2C9 contribute to the oxidative metabolism of voriconazole. When cytochrome P450 (P450)-specific inhibitors and antibodies were used to evaluate the oxidative metabolism of voriconazole by human liver microsomes, the results suggested that P450-mediated metabolism accounted for approximately 75% of the total oxidative metabolism. The studies presented here provide evidence that the remaining approximately 25% of the metabolic transformations are catalyzed by flavin-containing monooxygenase (FMO). This conclusion was based on the evidence that the NADPH-dependent metabolism of voriconazole was sensitive to heat (45 degrees C for 5 min), a condition known to selectively inactivate FMO without affecting P450 activity. The role of FMO in the metabolic formation of voriconazole N-oxide was confirmed by the use of recombinant FMO enzymes. Kinetic analysis of voriconazole metabolism by FMO1 and FMO3 yielded K(m) values of 3.0 and 3.4 mM and V(max) values of 0.025 and 0.044 pmol/min/pmol, respectively. FMO5 did not metabolize voriconazole effectively. This is the first report of the role of FMO in the oxidative metabolism of voriconazole.
Figures
References
-
- Allerston CK, Shimizu M, Fujieda M, Shephard EA, Yamazaki H, Phillips IR. Molecular evolution and balancing selection in the flavin-containing monooxygenase 3 gene (FMO3) Pharmacogenet Genomics. 2007;17:827–839. - PubMed
-
- Boucher HW, Groll AH, Chiou CC, Walsh TJ. Newer systemic antifungal agents : pharmacokinetics, safety and efficacy. Drug. 2004;64:1997–2020. - PubMed
-
- Cashman JR. Structural and catalytic properties of mammalian flavin-containing monooxygenase. Chem Res Toxicol. 1995;8:165–181. - PubMed
-
- Fisher MB, Yoon K, Vaughn ML, Strelevitz TJ, Foti RS. Flavin-containing monooxygenase activity in hepatocytes and microsomes: in vitro characterization and in vivo scaling of benzydamine clearance. Drug Metab Dispos. 2002;30:1087–1093. - PubMed
-
- Hines RN. Developmental and tissue-specific expression of human flavin-containing monooxygenases 1 and 3. Expert Opin Drug Metab Toxicol. 2006;2:41–49. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
