

Apoptosis induces expression of sphingosine kinase 1 to release sphingosine-1-phosphate as a "come-and-get-me" signal
- PMID: 18362204
- PMCID: PMC2493451
- DOI: 10.1096/fj.08-107169
Apoptosis induces expression of sphingosine kinase 1 to release sphingosine-1-phosphate as a "come-and-get-me" signal
Abstract
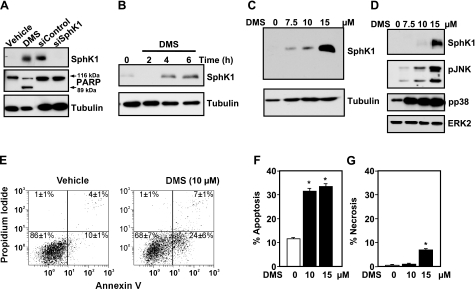
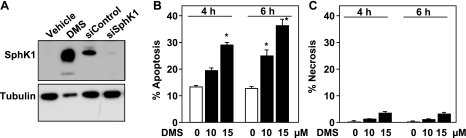
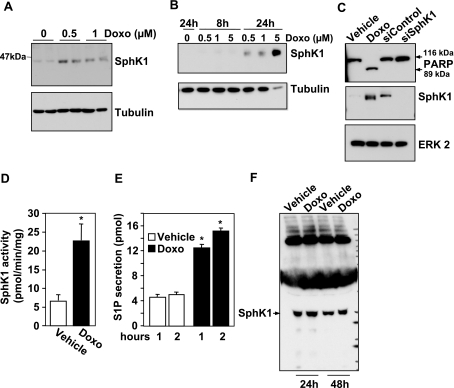
Sphingosine-1-phosphate (S1P) is a bioactive lipid that regulates myriad important cellular processes, including growth, survival, cytoskeleton rearrangements, motility, and immunity. Here we report that treatment of Jurkat and U937 leukemia cells with the pan-sphingosine kinase (SphK) inhibitor N,N-dimethylsphingosine to block S1P formation surprisingly caused a large increase in expression of SphK1 concomitant with induction of apoptosis. Another SphK inhibitor, D,L-threo-dihydrosphingosine, also induced apoptosis and produced dramatic increases in SphK1 expression. However, up-regulation of SphK1 was not a specific effect of its inhibition but rather was a consequence of apoptotic stress. The chemotherapeutic drug doxorubicin, a potent inducer of apoptosis in these cells, also stimulated SphK1 expression and activity and promoted S1P secretion. The caspase inhibitor ZVAD reduced not only doxorubicin-induced lethality but also the increased expression of SphK1 and secretion of S1P. Apoptotic cells secrete chemotactic factors to attract phagocytic cells, and we found that S1P potently stimulated chemotaxis of monocytic THP-1 and U937 cells and primary monocytes and macrophages. Collectively, our data suggest that apoptotic cells may up-regulate SphK1 to produce and secrete S1P that serves as a "come-and-get-me" signal for scavenger cells to engulf them in order to prevent necrosis.
Figures



References
-
- Spiegel S, Milstien S. Sphingosine-1-phosphate: an enigmatic signalling lipid. Nat Rev Mol Cell Biol. 2003;4:397–407. - PubMed
-
- Ogretmen B, Hannun Y A. Biologically active sphingolipids in cancer pathogenesis and treatment. Nat Rev Cancer. 2004;4:604–616. - PubMed
-
- Cuvillier O, Pirianov G, Kleuser B, Vanek P G, Coso O A, Gutkind S, Spiegel S. Suppression of ceramide-mediated programmed cell death by sphingosine-1-phosphate. Nature. 1996;381:800–803. - PubMed
-
- Taha T A, Hannun Y A, Obeid L M. Sphingosine kinase: biochemical and cellular regulation and role in disease. J Biochem Mol Biol. 2006;39:113–131. - PubMed
-
- Huwiler A, Pfeilschifter J. Altering the sphingosine-1-phosphate/ceramide balance: a promising approach for tumor therapy. Curr Pharm Des. 2006;12:4625–4635. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
