

IL-15 as a mediator of CD4+ help for CD8+ T cell longevity and avoidance of TRAIL-mediated apoptosis
- PMID: 18362335
- PMCID: PMC2278231
- DOI: 10.1073/pnas.0801003105
IL-15 as a mediator of CD4+ help for CD8+ T cell longevity and avoidance of TRAIL-mediated apoptosis
Abstract
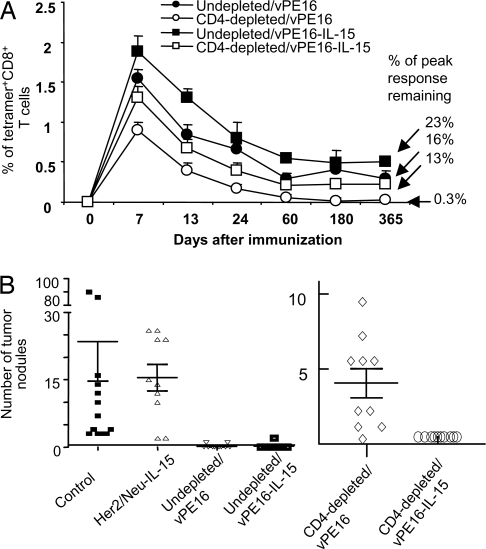
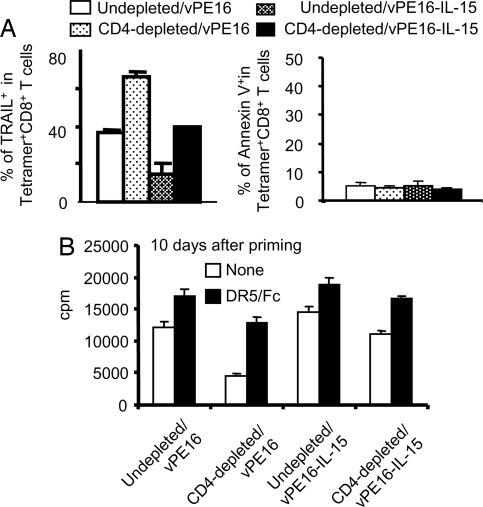
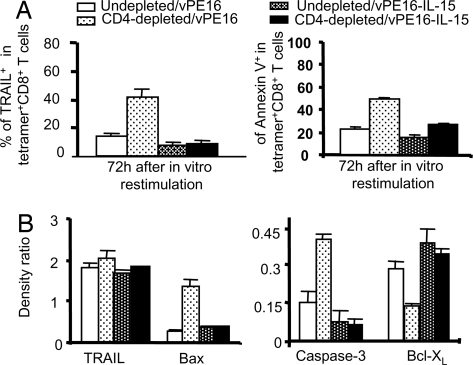
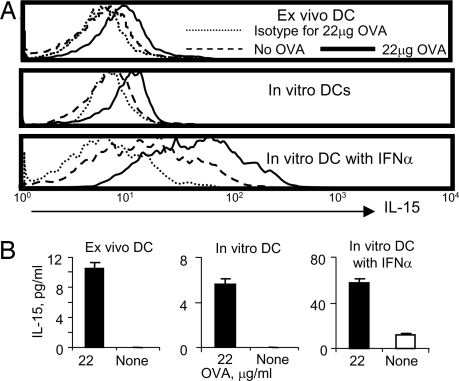
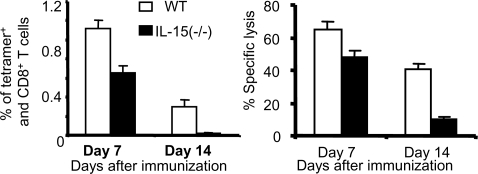
CD4+ helper T cells contribute to the induction and maintenance of antigen-specific CD8+ T cells. Their absence results in short-lived antigen-specific CD8+ T cells and defective secondary CD8+ T cell responses because of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-mediated apoptosis. Here, we show that IL-15 codelivered with vaccines can overcome CD4+ T cell deficiency for promoting longevity of antigen-specific CD8+ T cells and avoidance of TRAIL-mediated apoptosis. In both priming and secondary responses, IL-15 down-regulates proapoptotic Bax, an intermediate in TRAIL-mediated apoptosis, and increases anti-apoptotic Bcl-X(L) in CD8+ T cells. Thus, IL-15 is sufficient to mimic CD4+ T cell help. Antigen-specific CD4+ T cells induce dendritic cells (DCs) to produce IL-15. IL-15 is also necessary for optimal help, because helper cells do not deliver effective help through IL-15-/- DCs. Therefore, IL-15 codelivered with vaccines can overcome CD4+ helper T cell deficiency for induction of functionally efficient CD8+ T cells and maintenance of CD8+ cytotoxic T lymphocytes (CTLs), and IL-15 is probably one of the natural mediators of help. These findings suggest new vaccine strategies against infections and cancers, especially in individuals with CD4-deficiency.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Sprent J, Surh CD. T cell memory. Annu Rev Immunol. 2002;20:551–579. - PubMed
-
- Kaech SM, Wherry EJ, Ahmed R. Effector and memory T-cell differentiation: Implications for vaccine development. Nat Rev Immunol. 2002;2:251–262. - PubMed
-
- Badovinac VP, Porter BB, Harty JT. Programmed contraction of CD8+ T cells after infection. Nat Immunol. 2002;3:619–626. - PubMed
-
- Van Stipdonk MJ, et al. Dynamic programming of CD8+ T lymphocyte responses. Nat Immunol. 2003;4:361–365. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
