

The editing enzyme ADAR1 and the mRNA surveillance protein hUpf1 interact in the cell nucleus
- PMID: 18362360
- PMCID: PMC2278206
- DOI: 10.1073/pnas.0710576105
The editing enzyme ADAR1 and the mRNA surveillance protein hUpf1 interact in the cell nucleus
Abstract
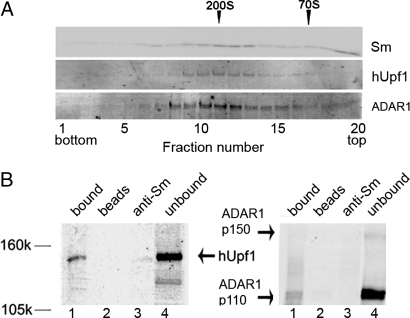
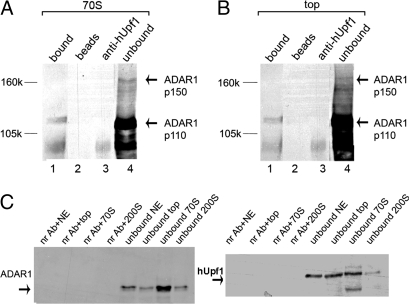
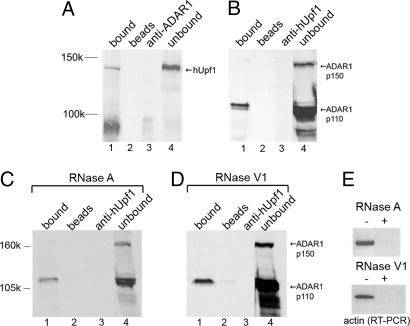
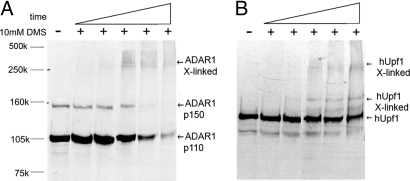
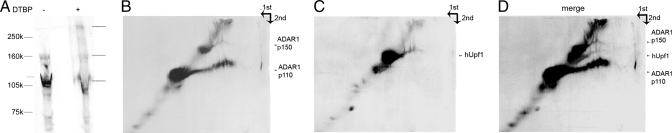
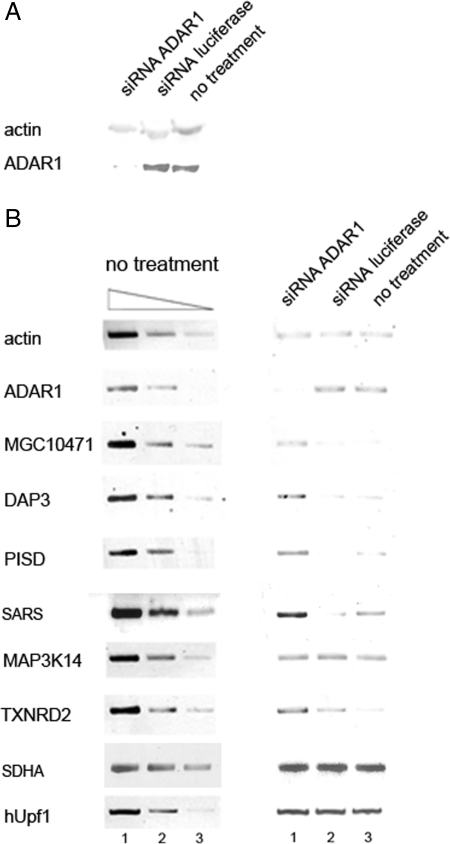
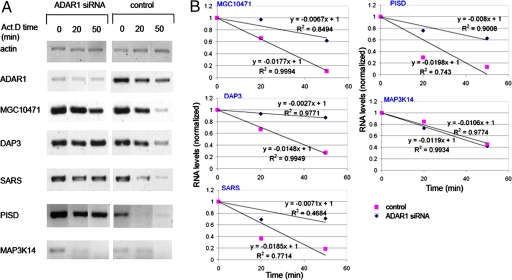
Posttranscriptional regulation is an important step in the regulation of gene expression. In this article, we show an unexpected connection between two proteins that participate in different processes of posttranscriptional regulation that ensures the production of functional mRNA molecules. Specifically, we show that the A-to-I RNA editing protein adenosine deaminase that acts on RNA 1 (ADAR1) and the human Upf1 (hUpf1) protein involved in RNA surveillance are found associated within nuclear RNA-splicing complexes. A potential functional role for this association was revealed by RNAi-mediated down-regulation of ADAR1, which was accompanied by up-regulation of a number of genes previously shown to undergo A-to-I editing in Alu repeats and to be down-regulated by hUpf1. This study suggests a regulatory pathway by a combination of ADAR1 A-to-I editing enzyme and RNA degradation presumably with the aid of hUpf1.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Valente L, Nishikura K. ADAR gene family and A-to-I RNA editing: Diverse roles in posttranscriptional gene regulation. Prog Nucleic Acid Res Mol Biol. 2005;79:299–338. - PubMed
-
- Sommer B, Kohler M, Sprengel R, Seeburg PH. RNA editing in brain controls a determinant of ion flow in glutamate-gated channels. Cell. 1991;67:11–19. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
