

Structural and biophysical characterization of human myo-inositol oxygenase
- PMID: 18364358
- PMCID: PMC3258897
- DOI: 10.1074/jbc.M800348200
Structural and biophysical characterization of human myo-inositol oxygenase
Abstract
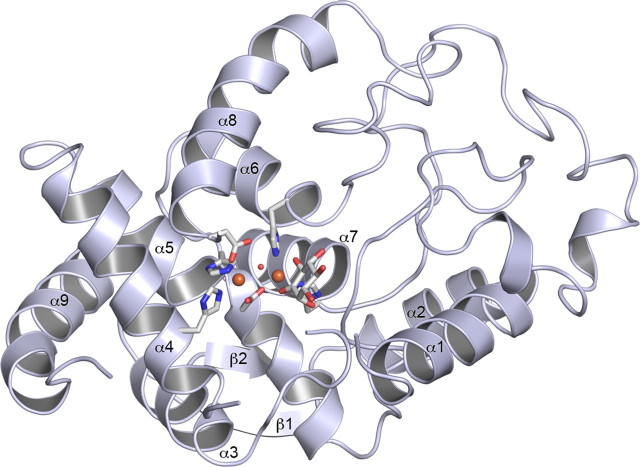
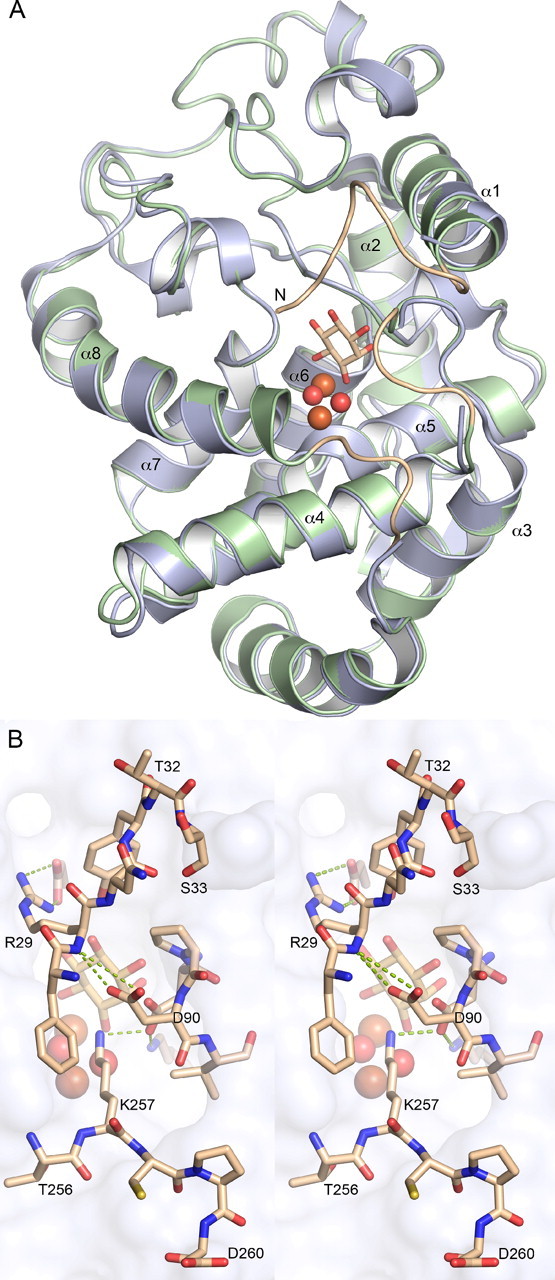
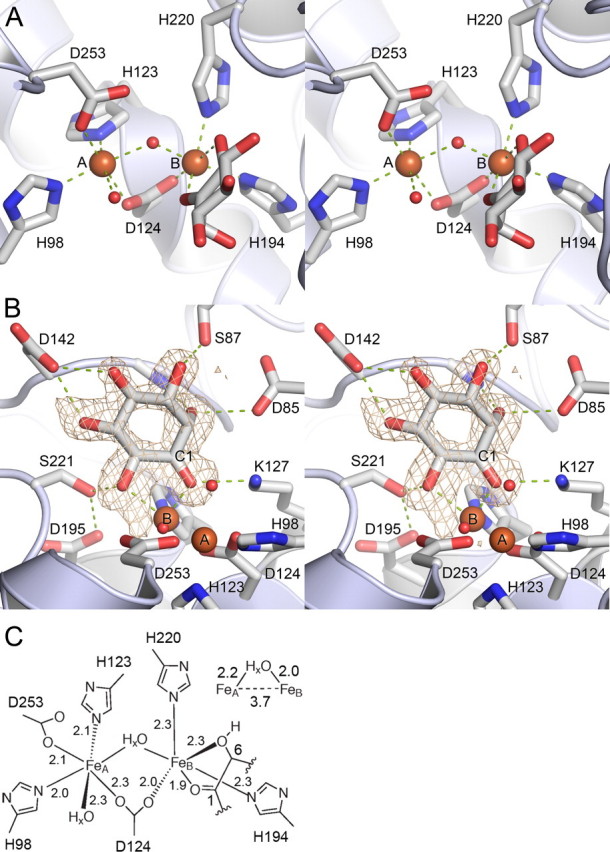
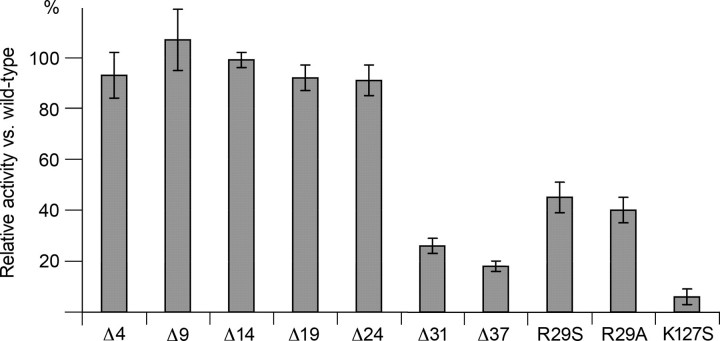
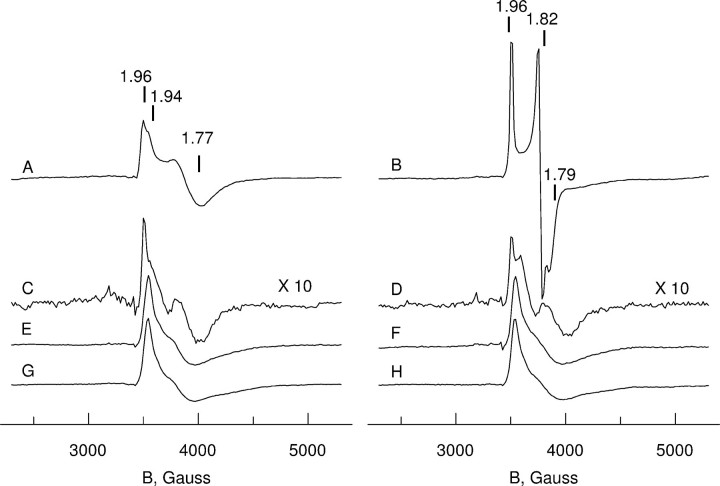
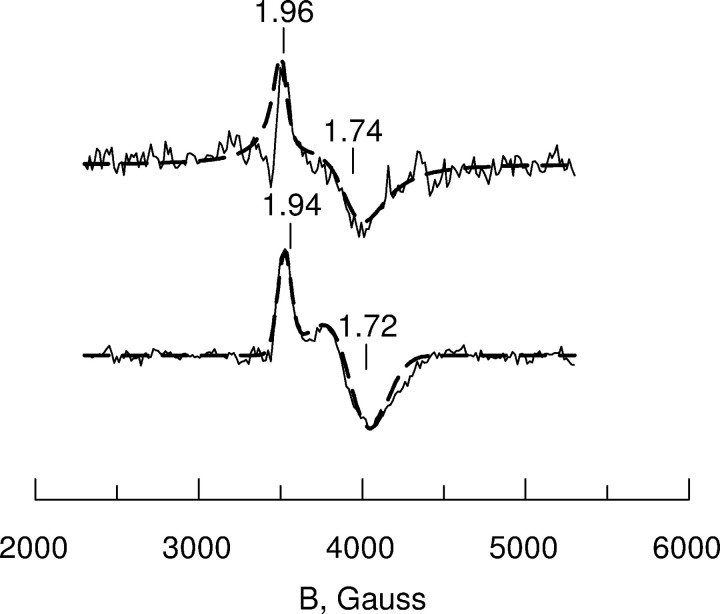
Altered inositol metabolism is implicated in a number of diabetic complications. The first committed step in mammalian inositol catabolism is performed by myo-inositol oxygenase (MIOX), which catalyzes a unique four-electron dioxygen-dependent ring cleavage of myo-inositol to D-glucuronate. Here, we present the crystal structure of human MIOX in complex with myo-inosose-1 bound in a terminal mode to the MIOX diiron cluster site. Furthermore, from biochemical and biophysical results from N-terminal deletion mutagenesis we show that the N terminus is important, through coordination of a set of loops covering the active site, in shielding the active site during catalysis. EPR spectroscopy of the unliganded enzyme displays a two-component spectrum that we can relate to an open and a closed active site conformation. Furthermore, based on site-directed mutagenesis in combination with biochemical and biophysical data, we propose a novel role for Lys(127) in governing access to the diiron cluster.
Figures
References
-
- Winegrad, A. I. (1987) Diabetes 36 396-406 - PubMed
-
- Naber, N. I., Swan, J. S., and Hamilton, G. A. (1986) Biochemistry 25 7201-7207 - PubMed
-
- Arner, R. J., Prabhu, K. S., and Reddy, C. C. (2004) Biochem. Biophys. Res. Commun. 324 1386-1392 - PubMed
-
- Arner, R. J., Prabhu, K. S., Krishnan, V., Johnson, M. C., and Reddy, C. C. (2006) Biochem. Biophys. Res. Commun. 339 816-820 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
