

Exclusion of a proton ATPase from the apical membrane is associated with cell polarity and tip growth in Nicotiana tabacum pollen tubes
- PMID: 18364468
- PMCID: PMC2329945
- DOI: 10.1105/tpc.106.047423
Exclusion of a proton ATPase from the apical membrane is associated with cell polarity and tip growth in Nicotiana tabacum pollen tubes
Abstract
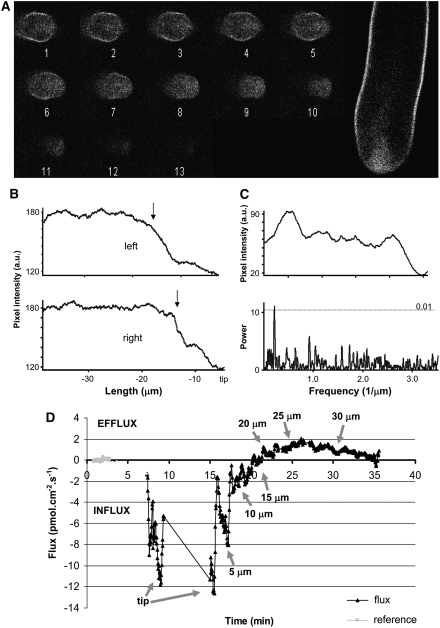
Polarized growth in pollen tubes results from exocytosis at the tip and is associated with conspicuous polarization of Ca(2+), H(+), K(+), and Cl(-) -fluxes. Here, we show that cell polarity in Nicotiana tabacum pollen is associated with the exclusion of a novel pollen-specific H(+)-ATPase, Nt AHA, from the growing apex. Nt AHA colocalizes with extracellular H(+) effluxes, which revert to influxes where Nt AHA is absent. Fluorescence recovery after photobleaching analysis showed that Nt AHA moves toward the apex of growing pollen tubes, suggesting that the major mechanism of insertion is not through apical exocytosis. Nt AHA mRNA is also excluded from the tip, suggesting a mechanism of polarization acting at the level of translation. Localized applications of the cation ionophore gramicidin A had no effect where Nt AHA was present but acidified the cytosol and induced reorientation of the pollen tube where Nt AHA was absent. Transgenic pollen overexpressing Nt AHA-GFP developed abnormal callose plugs accompanied by abnormal H(+) flux profiles. Furthermore, there is no net flux of H(+) in defined patches of membrane where callose plugs are to be formed. Taken together, our results suggest that proton dynamics may underlie basic mechanisms of polarity and spatial regulation in growing pollen tubes.
Figures







Similar articles
-
Pollen tube tip growth depends on plasma membrane polarization mediated by tobacco PLC3 activity and endocytic membrane recycling.Plant Cell. 2006 Dec;18(12):3519-34. doi: 10.1105/tpc.106.047373. Epub 2006 Dec 15. Plant Cell. 2006. PMID: 17172355 Free PMC article.
-
Localization of Arabidopsis SYP125 syntaxin in the plasma membrane sub-apical and distal zones of growing pollen tubes.Plant Signal Behav. 2011 May;6(5):665-70. doi: 10.4161/psb.6.5.14423. Epub 2011 May 1. Plant Signal Behav. 2011. PMID: 21499026 Free PMC article.
-
Type B phosphatidylinositol-4-phosphate 5-kinases mediate Arabidopsis and Nicotiana tabacum pollen tube growth by regulating apical pectin secretion.Plant Cell. 2008 Dec;20(12):3312-30. doi: 10.1105/tpc.108.059568. Epub 2008 Dec 5. Plant Cell. 2008. PMID: 19060112 Free PMC article.
-
The role of ion fluxes in polarized cell growth and morphogenesis: the pollen tube as an experimental paradigm.Int J Dev Biol. 2009;53(8-10):1609-22. doi: 10.1387/ijdb.072296em. Int J Dev Biol. 2009. PMID: 19247955 Review.
-
Rapid tip growth: insights from pollen tubes.Semin Cell Dev Biol. 2011 Oct;22(8):816-24. doi: 10.1016/j.semcdb.2011.06.004. Epub 2011 Jun 25. Semin Cell Dev Biol. 2011. PMID: 21729760 Free PMC article. Review.
Cited by
-
Perspectives for using genetically encoded fluorescent biosensors in plants.Front Plant Sci. 2013 Jul 12;4:234. doi: 10.3389/fpls.2013.00234. eCollection 2013. Front Plant Sci. 2013. PMID: 23874345 Free PMC article.
-
Control of cell wall extensibility during pollen tube growth.Mol Plant. 2013 Jul;6(4):998-1017. doi: 10.1093/mp/sst103. Epub 2013 Jun 14. Mol Plant. 2013. PMID: 23770837 Free PMC article. Review.
-
Arabidopsis LIM proteins: a family of actin bundlers with distinct expression patterns and modes of regulation.Plant Cell. 2010 Sep;22(9):3034-52. doi: 10.1105/tpc.110.075960. Epub 2010 Sep 3. Plant Cell. 2010. PMID: 20817848 Free PMC article.
-
Osmoregulation in Lilium pollen grains occurs via modulation of the plasma membrane H+ ATPase activity by 14-3-3 proteins.Plant Physiol. 2010 Dec;154(4):1921-8. doi: 10.1104/pp.110.165696. Epub 2010 Oct 25. Plant Physiol. 2010. PMID: 20974894 Free PMC article.
-
The transcription factor PbrbZIP52 positively affects pear pollen tube longevity by promoting callose synthesis.Plant Physiol. 2023 Mar 17;191(3):1734-1750. doi: 10.1093/plphys/kiad002. Plant Physiol. 2023. PMID: 36617219 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
