

Crystal structure of human spermine synthase: implications of substrate binding and catalytic mechanism
- PMID: 18367445
- PMCID: PMC3259631
- DOI: 10.1074/jbc.M710323200
Crystal structure of human spermine synthase: implications of substrate binding and catalytic mechanism
Abstract
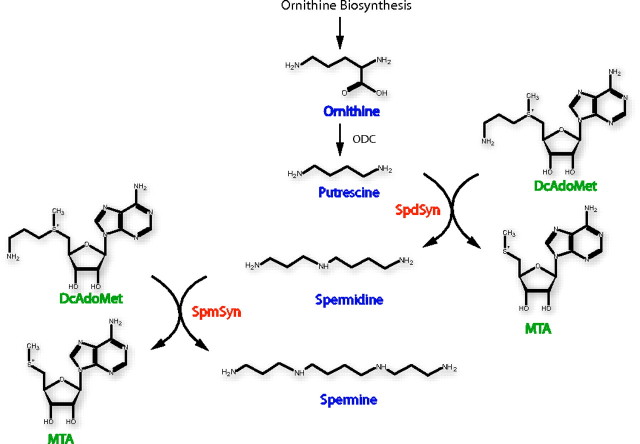
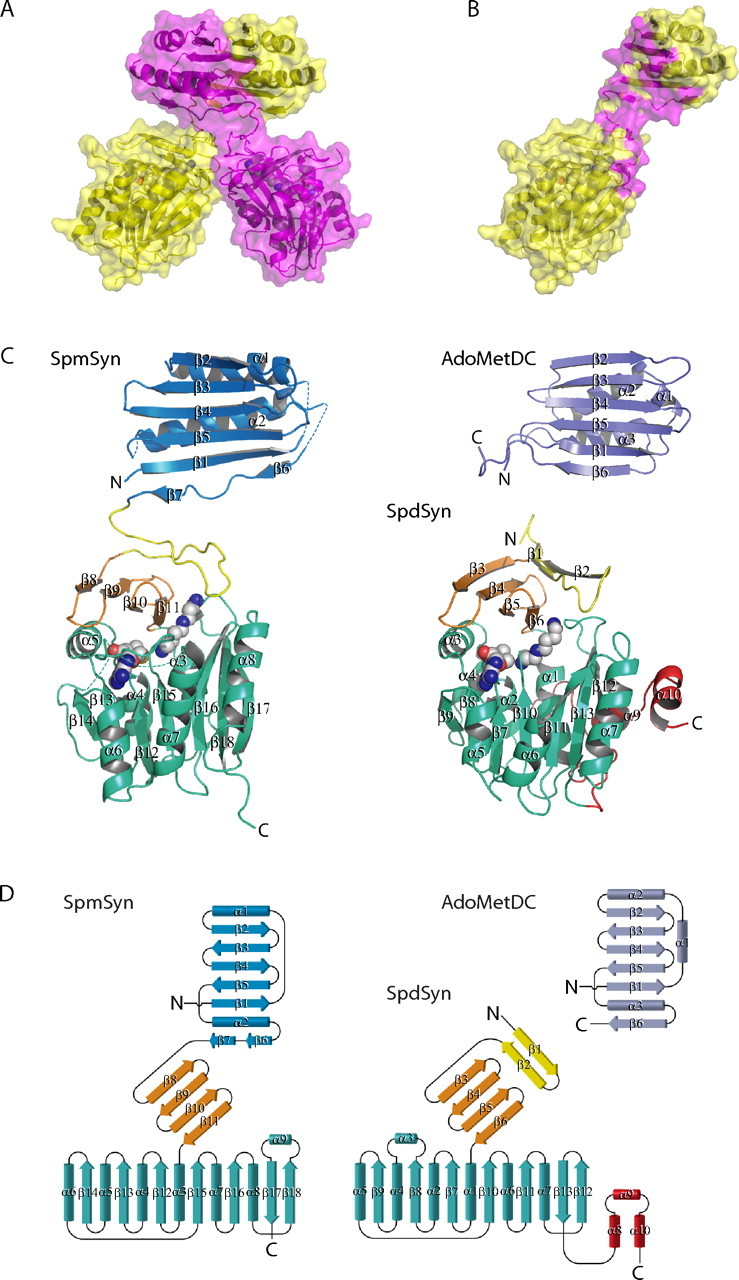
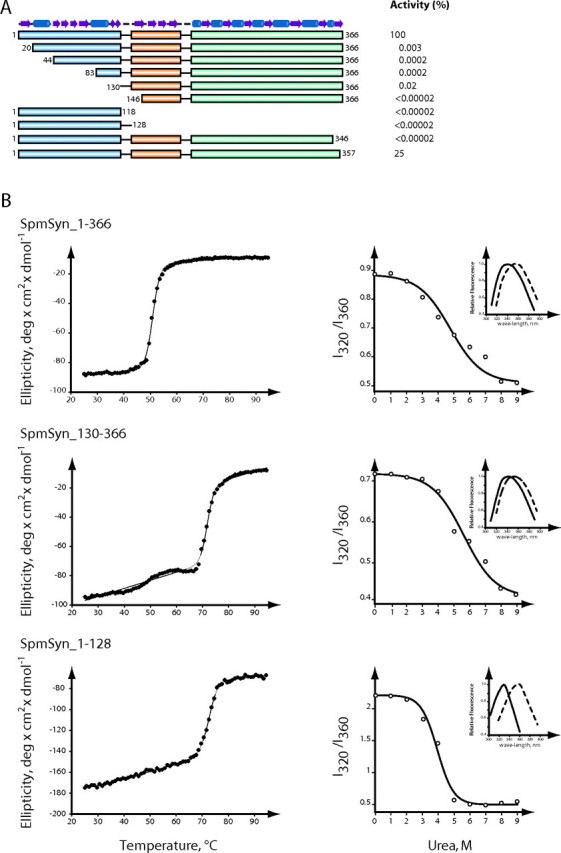
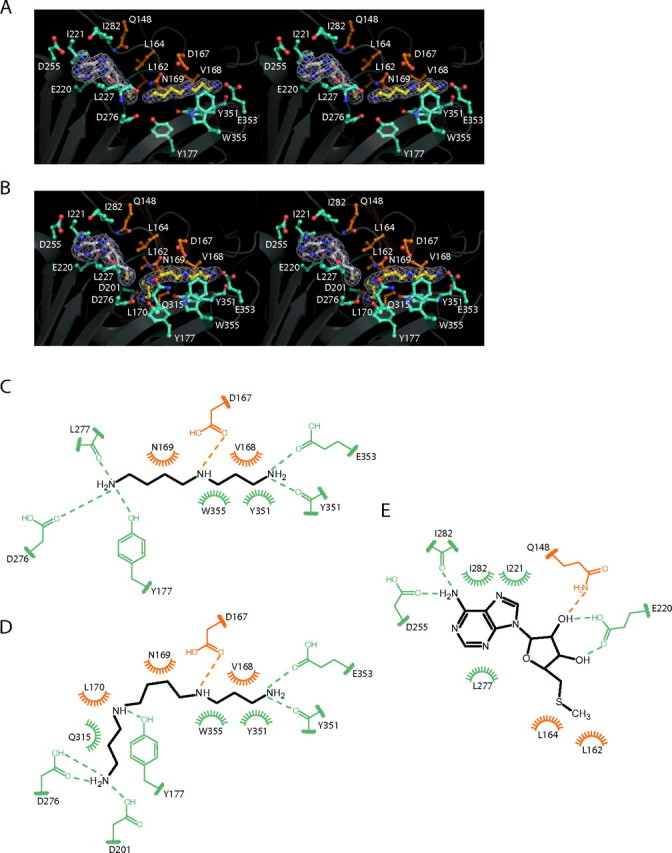
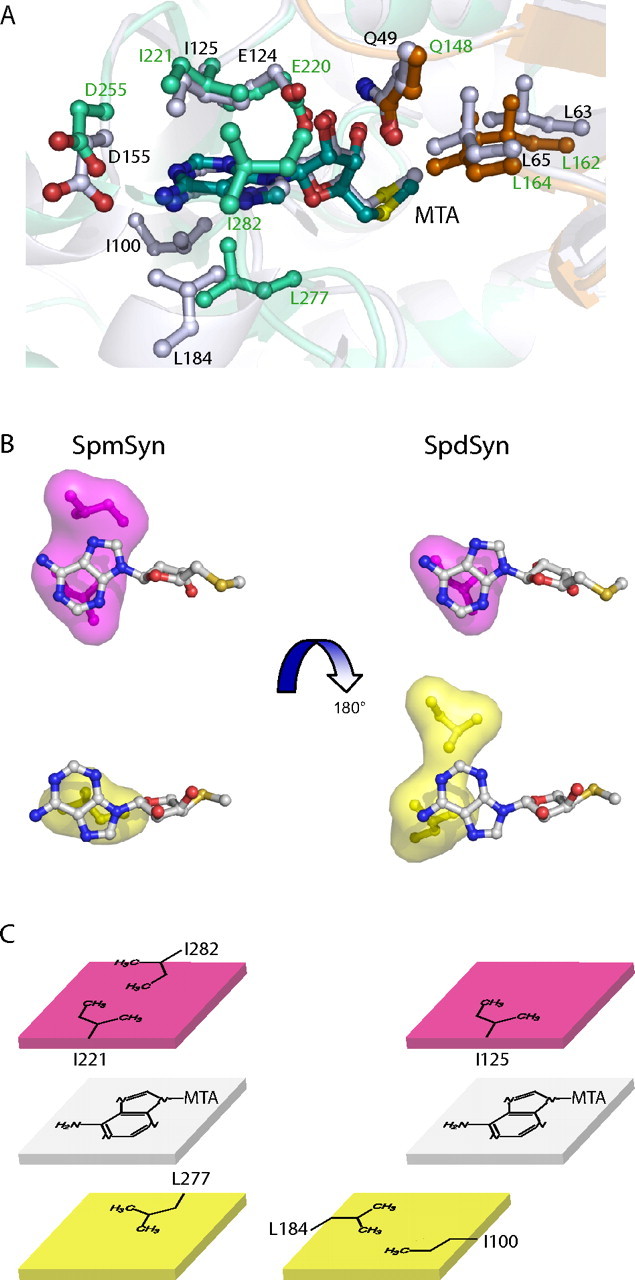
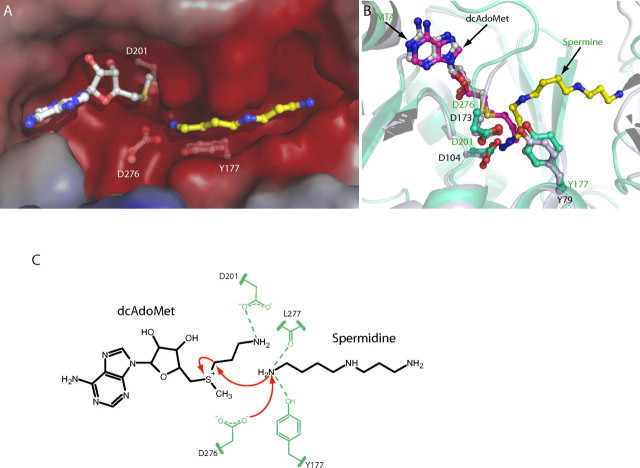
The crystal structures of two ternary complexes of human spermine synthase (EC 2.5.1.22), one with 5'-methylthioadenosine and spermidine and the other with 5'-methylthioadenosine and spermine, have been solved. They show that the enzyme is a dimer of two identical subunits. Each monomer has three domains: a C-terminal domain, which contains the active site and is similar in structure to spermidine synthase; a central domain made up of four beta-strands; and an N-terminal domain with remarkable structural similarity to S-adenosylmethionine decarboxylase, the enzyme that forms the aminopropyl donor substrate. Dimerization occurs mainly through interactions between the N-terminal domains. Deletion of the N-terminal domain led to a complete loss of spermine synthase activity, suggesting that dimerization may be required for activity. The structures provide an outline of the active site and a plausible model for catalysis. The active site is similar to those of spermidine synthases but has a larger substrate-binding pocket able to accommodate longer substrates. Two residues (Asp(201) and Asp(276)) that are conserved in aminopropyltransferases appear to play a key part in the catalytic mechanism, and this role was supported by the results of site-directed mutagenesis. The spermine synthase.5'-methylthioadenosine structure provides a plausible explanation for the potent inhibition of the reaction by this product and the stronger inhibition of spermine synthase compared with spermidine synthase. An analysis to trace possible evolutionary origins of spermine synthase is also described.
Figures


References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
